| The
Microbial World Microorganisms and microbial activities Produced by
Jim Deacon |
||
This site is no longer maintained and has been left for archival purposes
Text and links may be out of date
| The
Microbial World Microorganisms and microbial activities Produced by
Jim Deacon |
||
I welcome comments and suggestions at: j.deacon@ed.ac.uk DISCLAIMER. The information in these pages is believed to be accurate, but the author provides no warranty to this effect and will not be liable for the consequence(s) of any reliance placed on the information. |
|
|
This is a clickable
image |
| The microbial world.
I. Activities of microorganisms Microorganisms are everywhere - a largely unseen world of activities that helped to create the biosphere and that continue to support the life processes on earth. So, Welcome to the Microbial World. In fact, you are part of it! Of all the cells that make up the normal, healthy human body, more than 99 per cent are the cells of microorganisms living on the skin or in the gut, etc. This normal resident microbial population includes potential pathogens as well as organisms that help to keep the potential pathogens in check (see Yeasts and yeast-like fungi). Here are a few more examples of the impact of microorganisms.
|
||
The main plants seen here are: the giant saguaro cactus (Carnegiea gigantea, about 6 metres tall), various species of Opuntia (large jointed pads of the prickly pear to the left and right of the saguaro, teddy bear cholla in the foreground, the purple coloured pencil cholla to the left of the saguaro), bursage (Ambrosia deltoidea) in much of the foreground, and paloverde (Parkinsonia), the green-stemmed tree to the right of the saguaro. [Image supplied by Sharon von Broembsen, Oklahoma State University] |
||
Our list could go on, but we end this section by noting the astonishing diversity of physiology and metabolism that enables microbes to grow in every environment that will support life on earth.
|
||
Continue on this page for: The Microbial World. II. Origins and Diversity
|
||
| The Microbial World. II. Origins
and Diversity Here we consider microorganisms in geological time - how they evolved and diversified, and how they created the world that we know today. We also see the place of microorganisms in the scheme of life as a whole, based on gene sequencing to analyse the relatedness of organisms. |
||||||||||||||||||||||||
| History of life on earth An approximate history of life on earth is shown in the table below, based on information in Encyclopaedia Britannica (1986). There may be more up-to-date information, but the major points would still apply. The evidence is fragmentary, and is obtained from three major sources:
|
||||||||||||||||||||||||
From this table we see several key points.
|
||||||||||||||||||||||||
| Microorganisms and
the "Tree of Life" Biologists have always striven to find a universal "tree of life" - a tree that reflects the natural, evolutionary relationships of living organisms and that, hopefully, extends back to the very origins of life. Prokaryotes and eukaryotes An important step along the path was the recognition that living organisms can be separated into two basic types: those with cells that contain a nucleus (the eukaryotes) and those that lack a nucleus (prokaryotes). Essentially, all the multicellular life forms are eukaryotes (with larger cells, containing organelles and a relatively large genome distributed between several chromosomes) whereas all bacteria and bacterium-like organisms are prokaryotes (with small cells, no internal membrane-bound organelles and a single circular chromosome). The geological record shows that organisms resembling today's prokaryotes have existed on earth for probably 3,500 million years, whereas eukaryotes have existed for perhaps only 1,500-2,000 million years. The "Five Kingdoms" In recent years biologists have tended to recognise five Kingdoms of organisms: the Monera (bacteria and bacterium-like organisms) representing prokaryotes, and plants, animals, fungi and protists (mainly unicellular nucleate organisms) representing eukaryotes. The Five Kingdom approach is attractive in its simplicity, but has significant problems. One of these concerns the protists - a wide range of disparate organisms such as amoebae, slime moulds, ciliates, algae, etc. that are grouped together as a kingdom with little justification. Another problem stems from the recognition in the 1980s that some bacterium-like organisms (first given the name archaebacteria, and now called archaea) are so different from the true bacteria that they can be separated as a group. They are prokaryotes, and they look like bacteria, but in terms of cellular biochemistry and genetics the archaea differ from both eukaryotes and bacteria (see below). DNA sequencing DNA sequencing has provided a new approach for studying evolutionary relationships, since:
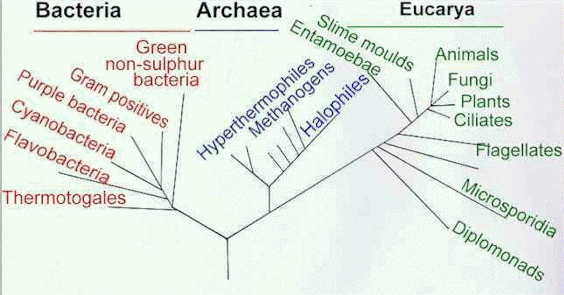
So, by comparing the genes that code for vital functions of all living organisms, it should be possible to assess the relatedness of different organisms. The gene most commonly used for this codes for the RNA in the small subunit (SSU) of the ribosome. [Ribosomes are the structures on which proteins are synthesised]. Some regions of this SSU rRNA (also termed 16S rRNA) are highly conserved in all organisms, whereas other regions are more variable. By comparing the DNA sequences for 16S rRNA, Woese and his colleagues constructed a proposed universal phylogenetic tree, shown in simplified form below. |
||||||||||||||||||||||||
Proposed universal phylogenetic tree, based on nucleotide sequence comparisons of the DNA coding for the RNA of the small ribosomal subunit of different organisms. Only some of the microbial groups are shown here. [Based on a diagram in R Woese (1994) Microbiological Reviews 58, 1-9.] |
||||||||||||||||||||||||
We should note a technical point about this tree: the comparison of ribosomal RNA gene sequences can show the possible relatedness of organisms, but other information is needed to provide the root of a tree. One of the principal modes of evolution is thought to involve gene duplication followed by divergence. The original gene retains its vital function, while the copy can change and ultimately can encode a new function. If these paralogous gene pairs can be identified by sequence similarity, then the original gene should be present in all organisms whereas the new version will be present only in the more recently derived organisms. The root for the tree in the diagram above was determined by using paralogous genes for translation elongation factors involved in synthesis of protein chains on the ribosomes. Domains and Kingdoms The proposed universal phylogenetic tree recognises three Domains of organisms (Bacteria, Archaea and Eucarya) above the traditional level of Kingdoms. These domains seem to have diverged from one another a long time ago, presumably from an extinct or as yet undiscovered ancestral line. As shown in the diagram above, the archaea and eucarya seem to have arisen from a common line more recently than the divergence of these two groups from the bacteria. However, we should note that this tree is only provisional - it can look quite different if it is rooted by other methods. There will be many changes before we arrive at a universally acceptable tree, if ever!
What should we now recognise as Kingdoms? Perhaps all the terminal branches of the tree represent kingdoms or taxa of similar major rank, and more will be added as the sequences of other organisms become available. Clearly, some of these kingdoms (or potential kingdoms) have advanced and expanded much more than others - the plants, animals and fungi are major groups of organisms with distinctive lifestyles, whereas the slime moulds and ciliates, for example, have not expanded to the same degree. Origin and evolution of eukaryotes Eukaryotes must have arisen from prokaryotic ancestors. Many aspects of this are still unknown, but there is persuasive evidence that the mitochondria and plastids (chloroplasts) of today’s eukaryotes are derived from prokaryotes. For example, both mitochondria and plastids have a single circle of DNA (the vestige of a bacterial chromosome) which codes for some of their functions. Also, mitochondria and plastids have ribosomes that resemble those of prokaryotes (termed 70S ribosomes) and that are sensitive to antibacterial antibiotics. Gene sequencing shows that:
Thus, it is believed that these organelles of eukaryotes represent bacteria that once lived inside the cells of other bacteria. Over evolutionary time, the endosymbionts (ntermal dwellers) lost the ability for an independent life and became reduced to the state of serving particular functions (oxidative energy metabolism, and photosynthesis) in eukaryotes. It is much more difficult to trace the origins of two other characteristic features of eukaryotes - the nucleus and the cytoskeleton (which is composed or microtubules and associated proteins). Lateral gene transfer Genealogies based on the genes coding for small subunit ribosomal RNA (as in the diagram above) provide a rather simple view of an ordered sequence of evolutionary progression. However, the situation becomes more complex when these findings are supplemented with the DNA sequence data for other genes, such as those encoding important enzyme functions. Then it becomes clear that the genomes of eukaryotes are chimeric - they have some gene sequences that clearly resemble those of the archaea and some that resemble those of bacteria. In other words, there does not seem to be an ordered, linear sequence of genetic change during the early evolution of the major groups of eukaryotes. Instead, there is likely to have been one or more gene transfer events between the evolving lines of bacteria, archaea and eucarya. One way in which this could have happened is by engulfment of one organism by another (perhaps followed by endosymbiosis), then some of the engulfed organism's genes might have been retained while others were lost over time. For further discussion of all these points, see: Symposia of the Society for General Microbiology Volume 54. Evolution of Microbial Life (1996) Cambridge University Press. Laura A Katz (1998) Changing perspectives on the origin of eukaryotes. Trends in Ecology and Evolution Volume 13, 493-497. The archaea The archaea are a fascinating group of organisms. Although they look like bacteria, and have simple cells with a prokaryotic structure, they have biochemical and genetic features quite different from those of bacteria. For example:
In terms of physiology and behaviour, the archaea fall into three major types.
The growth conditions for the archaea are thought to resemble those that existed in the early stages of the earth's history. Thus, these organisms were termed archaebacteria (ancient bacteria) when they were first discovered. But we should not assume that they were necessarily the first microorganisms. Newly discovered microbes We have always known that the current methods of sampling and culturing of organisms from natural environments are deficient - these methods tend to select for the fastest-growing organisms in the culture conditions that are used. But the DNA sequencing methods discussed earlier have been a powerful new tool for detecting and ultimately isolating unknown (and even unsuspected) microorganisms. Basically, DNA is extracted from an environmental sample, split into single strands and mixed with a primer - a short DNA sequence that will combine with the complementary sequence on DNA in the sample. Then by the polymerase chain reaction the DNA in the environmental sample will be synthesised, starting at the primer and copying along the DNA strand. By using a primer from a highly conserved region of the gene for SSU rRNA, any ribosomal genes in the sample will be copied and their sequences can be compared with the sequences of known organisms. Any new gene sequences will represent new organisms, and these can be placed within the SSU rRNA gene tree. In this way, two new types of archaea have been discovered from marine environments in the last decade. They seem to be remarkably common around the world, and the abundance of their DNA in coastal surface waters suggests that they can represent about 34% of the prokaryote biomass at certain times of the year.
|
||||||||||||||||||||||||
Further reading: websites For more on the archaea and molecular evolution, some good starting points are: The Scientist (not on this server) The Tree of Life Project (not on this server)
|
About this site Aims and history
Contributors
|
Links to Microbiology and related websites
Fungal websites
Plant pathology websites
|
This site is no longer maintained and has been left for archival purposes
Text and links may be out of date