This site is no longer maintained and has been left for archival purposes
Text and links may be out of date
| Nitrogen fixation A relatively small amount of ammonia is produced by lightning. Some ammonia also is produced industrially by the Haber-Bosch process, using an iron-based catalyst, very high pressures and fairly high temperature. But the major conversion of N2 into ammonia, and thence into proteins, is achieved by microorganisms in the process called nitrogen fixation (or dinitrogen fixation). The table below shows some estimates of the amount of nitrogen fixed on a global scale. The total biological nitrogen fixation is estimated to be twice as much as the total nitrogen fixation by non-biological processes. |
|
||||||||||||||||||||||||||
| To illustrate the
importance of biological nitrogen fixation, the image
below shows part of the Lower Sonoran desert in Arizona.
Every plant that we see in this scene depends ultimately
on biological nitrogen fixation. Both free-living cyanobacteria and
the cyanobacterial associates of lichens initially contributed nitrogen to the soil
by forming a cryptobiotic crust. Now numerous leguminous
plants occur in this desert, with nitrogen-fixing Rhizobium
in their root nodules. Examples are the green-stemmed brush-like trees at
the right and left of the image (Parkinsonia
species, common name "paloverde"), and several
acacias and mesquites (not seen in this image).
|
|||||||||||||||||||||||||||||||
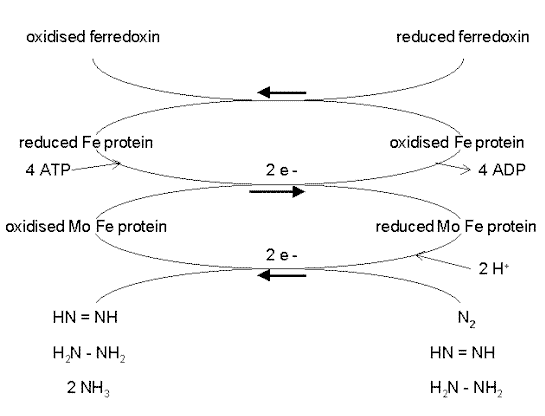
Mechanism of biological nitrogen fixation Biological nitrogen fixation can be represented by the following equation, in which two moles of ammonia are produced from one mole of nitrogen gas, at the expense of 16 moles of ATP and a supply of electrons and protons (hydrogen ions): N2 + 8H+ + 8e- + 16 ATP = 2NH3 + H2 + 16ADP + 16 Pi This reaction is performed exclusively by prokaryotes (the bacteria and related organisms), using an enzyme complex termed nitrogenase. This enzyme consists of two proteins - an iron protein and a molybdenum-iron protein, as shown below. The reactions occur while N2 is bound to the nitrogenase enzyme complex. The Fe protein is first reduced by electrons donated by ferredoxin. Then the reduced Fe protein binds ATP and reduces the molybdenum-iron protein, which donates electrons to N2, producing HN=NH. In two further cycles of this process (each requiring electrons donated by ferredoxin) HN=NH is reduced to H2N-NH2, and this in turn is reduced to 2NH3. Depending on the type of microorganism, the reduced ferredoxin which supplies electrons for this process is generated by photosynthesis, respiration or fermentation.
There is a remarkable degree of functional conservation between the nitrogenase proteins of all nitrogen-fixing bacteria. The Fe protein and the Mo-Fe protein have been isolated from many of these bacteria, and nitrogen fixation can be shown to occur in cell-free systems in a laboratory when the Fe protein of one species is mixed with the Mo-Fe protein of another bacterium, even if the species are very distantly related. |
|||||||||||||||||||||||||||||||
| The nitrogen-fixing organisms All the nitrogen-fixing organisms are
prokaryotes (bacteria). Some of them live independently
of other organisms - the so-called free-living
nitrogen-fixing bacteria. Others live in intimate
symbiotic associations with plants or with other
organisms (e.g. protozoa). Examples are shown in the
table below.
|
|||||||||||||||||||||||||||||||
| A point of special interest is that the nitrogenase enzyme complex is highly sensitive to oxygen. It is inactivated when exposed to oxygen, because this reacts with the iron component of the proteins. Although this is not a problem for anaerobic bacteria, it could be a major problem for the aerobic species such as cyanobacteria (which generate oxygen during photosynthesis) and the free-living aerobic bacteria of soils, such as Azotobacter and Beijerinckia. These organisms have various methods to overcome the problem. For example, Azotobacter species have the highest known rate of respiratory metabolism of any organism, so they might protect the enzyme by maintaining a very low level of oxygen in their cells. Azotobacter species also produce copious amounts of extracellular polysaccharide (as do Rhizobium species in culture - see Exopolysaccharides). By maintaining water within the polysaccharide slime layer, these bacteria can limit the diffusion rate of oxygen to the cells. In the symbiotic nitrogen-fixing organisms such as Rhizobium, the root nodules can contain oxygen-scavenging molecules such as leghaemoglobin, which shows as a pink colour when the active nitrogen-fixing nodules of legume roots are cut open. Leghaemoglobin may regulate the supply of oxygen to the nodule tissues in the same way as haemoglobin regulates the supply of oxygen to mammalian tissues. Some of the cyanobacteria have yet another mechanism for protecting nitrogenase: nitrogen fixation occurs in special cells (heterocysts) which possess only photosystem I (used to generate ATP by light-mediated reactions) whereas the other cells have both photosystem I and photosystem II (which generates oxygen when light energy is used to split water to supply H2 for synthesis of organic compounds). Extensive references to these points can be found at: http://chemweb.urich.edu/~sjanson/index.htm (not on this server) | |||||||||||||||||||||||||||||||
| Symbiotic nitrogen fixation 1. Legume symbioses The most familiar examples of nitrogen-fixing symbioses are the root nodules of legumes (peas, beans, clover, etc.).
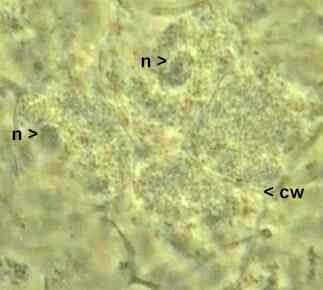
In these leguminous associations the bacteria usually are Rhizobium species, but the root nodules of soybeans, chickpea and some other legumes are formed by small-celled rhizobia termed Bradyrhizobium. Nodules on some tropical leguminous plants are formed by yet other genera. In all cases the bacteria "invade" the plant and cause the formation of a nodule by inducing localised proliferation of the plant host cells. Yet the bacteria always remain separated from the host cytoplasm by being enclosed in a membrane - a necessary feature in symbioses (see the image below).
Part of a crushed root nodule of a pea plant, showing four root cells containing colonies of Rhizobium. The nuclei (n) of two root cells are shown; cw indicates the cell wall that separates two plant cells. Although it cannot be seen clearly in this image, the bacteria occur in clusters which are enclosed in membranes, separating them from the cytoplasm of the plant cells. In nodules where nitrogen-fixation is occurring, the plant tissues contain the oxygen-scavenging molecule, leghaemoglobin (serving the same function as the oxygen-carrying haemoglobin in blood). The function of this molecule in nodules is to reduce the amount of free oxygen, and thereby to protect the nitrogen-fixing enzyme nitrogenase, which is irreversibly inactivated by oxygen. Some excellent images and discussion of these leguminous associations can be found at: CIFN/UNAM Brochure - Research (not on this server) |
|||||||||||||||||||||||||||||||
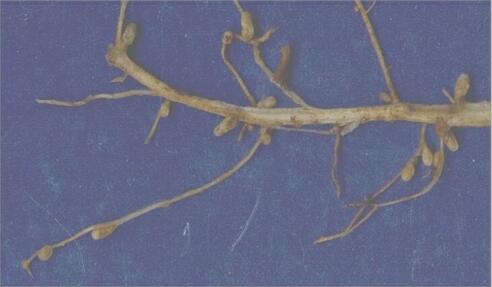
2. Associations with Frankia Frankia is a genus of the bacterial group termed actinomycetes - filamentous bacteria that are noted for their production of air-borne spores. Included in this group are the common soil-dwelling Streptomyces species which produce many of the antibiotics used in medicine (see Streptomyces). Frankia species are slow-growing in culture, and require specialised media, suggesting that they are specialised symbionts. They form nitrogen-fixing root nodules (sometimes called actinorhizae) with several woody plants of different families, such as alder (Alnus species), sea buckthorn (Hippophae rhamnoides, which is common in sand-dune environments) and Casuarina (a Mediterranean tree genus). Figure A (below) shows a young alder tree (Alnus glutinosa) growing in a plant pot, and Figure B shows part of the root system of this tree, bearing the orange-yellow coloured nodules (arrowheads) containing Frankia.
Alder and the other woody hosts of Frankia are typical pioneer species that invade nutrient-poor soils. These plants probably benefit from the nitrogen-fixing association, while supplying the bacterial symbiont with photosynthetic products. |
|||||||||||||||||||||||||||||||
| 3. Cyanobacterial associations The photosynthetic cyanobacteria often live as free-living organisms in pioneer habitats such as desert soils (see cyanobacteria) or as symbionts with lichens in other pioneer habitats. They also form symbiotic associations with other organisms such as the water fern Azolla, and cycads.The association with Azolla, where cyanobacteria (Anabaena azollae) are harboured in the leaves, has sometimes been shown to be important for nitrogen inputs in rice paddies, especially if the fern is allowed to grow and then ploughed into the soil to release nitrogen before the rice crop is sown. A symbiotic association of cyanobacteria with cycads is shown below. The first image shows a pot-grown plant. The second image shows a close-up of the soil surface in this pot. Short, club-shaped, branching roots have grown into the aerial environment. These aerial roots contain a nitrogen-fixing cyanobacterial symbiont.
|
|||||||||||||||||||||||||||||||
| In addition to these
intimate and specialised symbiotic associations, there
are several free-living nitrogen-fixing bacteria that
grow in close association with plants. For example, Azospirillum
species have been shown to fix nitrogen when growing in
the root zone (rhizosphere) or tropical grasses, and even
of maize plants in field conditions. Similarly, Azotobacter
species can fix nitrogen in the rhizosphere of several
plants. In both cases the bacteria grow at the expense of
sugars and other nutrients that leak from the roots.
However, these bacteria can make only a small
contribution to the nitrogen nutrition of the plant,
because nitrogen-fixation is an energy-expensive process,
and large amounts of organic nutrients are not
continuously available to microbes in the rhizosphere. This limitation may not apply to the bacteria that live in root nodules or other intimate symbiotic associations with plants. It has been estimated that nitrogen fixation in the nodules of clover roots or other leguminous plants may consume as much as 20% of the total photosynthate. |
|||||||||||||||||||||||||||||||
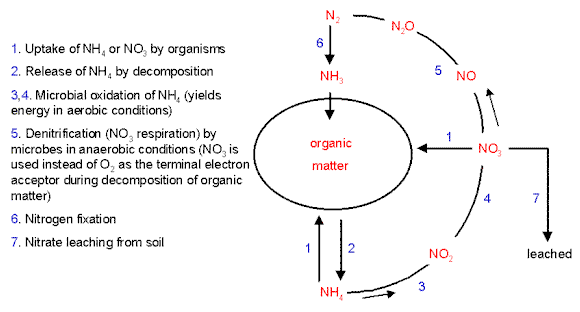
| The nitrogen cycle The diagram below shows an overview of the nitrogen cycle in soil or aquatic environments. At any one time a large proportion of the total fixed nitrogen will be locked up in the biomass or in the dead remains of organisms (shown collectively as "organic matter"). So, the only nitrogen available to support new growth will be that which is supplied by nitrogen fixation from the atmosphere (pathway 6 in the diagram) or by the release of ammonium or simple organic nitrogen compounds through the decomposition of organic matter (pathway 2). Some of other stages in this cycle are mediated by specialised groups of microorganisms and are explained below.
Nitrification The term nitrification refers to the conversion of ammonium to nitrate (pathway 3-4). This is brought about by the nitrifying bacteria, which are specialised to gain their energy by oxidising ammonium, while using CO2 as their source of carbon to synthesise organic compounds. Organisms of this sort are termed chemoautotrophs - they gain their energy by chemical oxidations (chemo-) and they are autotrophs (self-feeders) because they do not depend on pre-formed organic matter. In principle the oxidation of ammonium by these bacteria is no different from the way in which humans gain energy by oxidising sugars. Their use of CO2 to produce organic matter is no different in principle from the behaviour of plants. The nitrifying bacteria are found in most soils and waters of moderate pH, but are not active in highly acidic soils. They almost always are found as mixed-species communities (termed consortia) because some of them - e.g. Nitrosomonas species - are specialised to convert ammonium to nitrite (NO2-) while others - e.g. Nitrobacter species - convert nitrite to nitrate (NO3-). In fact, the accumulation of nitrite inhibits Nitrosomonas, so it depends on Nitrobacter to convert this to nitrate, whereas Nitrobacter depends on Nitrosomonas to generate nitrite. The nitrifying bacteria have some important environmental consequences, because they are so common that most of the ammonium in oxygenated soil or natural waters is readily converted to nitrate. Most plants and microorganisms can take up either nitrate or ammonium (arrows marked "1" in the diagram). However, process of nitrification has some undesirable consequences. The ammonium ion (NH4+) has a positive charge and so is readily adsorbed onto the negatively charged clay colloids and soil organic matter, preventing it from being washed out of the soil by rainfall. In contrast, the negatively charged nitrate ion is not held on soil particles and so can be washed down the soil profile - the process termed leaching (arrow marked 7 in the diagram). In this way, valuable nitrogen can be lost from the soil, reducing the soil fertility. The nitrates can then accumulate in groundwater, and ultimately in drinking water. There are strict regulations governing the amount of nitrate that can be present in drinking water, because nitrates can be reduced to highly reactive nitrites by microorganisms in the anaerobic conditions of the gut. Nitrites are absorbed from the gut and bind to haemoglobin, reducing its oxygen-carrying capacity. In young babies this can lead to respiratory distress - the condition known as "blue baby syndrome". Nitrite in the gut also can react with amino compounds, forming highly carcinogenic nitrosamines. Denitrification refers to the process in which nitrate is converted to gaseous compounds (nitric oxide, nitrous oxide and N2) by microorganisms. The sequence usually involves the production of nitrite (NO2-) as an intermediate step is shown as "5" in the diagram above. Several types of bacteria perform this conversion when growing on organic matter in anaerobic conditions. Because of the lack of oxygen for normal aerobic respiration, they use nitrate in place of oxygen as the terminal electron acceptor. This is termed anaerobic respiration and can be illustrated as follows: In aerobic respiration (as in humans), organic molecules are oxidised to obtain energy, while oxygen is reduced to water: C6H12O6 + 6 O2 = 6 CO2 + 6 H2O + energy In the absence of oxygen, any reducible substance such as nitrate (NO3-) could serve the same role and be reduced to nitrite, nitric oxide, nitrous oxide or N2. Thus, the conditions in which we find denitrifying organisms are characterised by (1) a supply of oxidisable organic matter, and (2) absence of oxygen but availability of reducible nitrogen sources. A mixture of gaseous nitrogen products is often produced because of the stepwise use of nitrate, nitrite, nitric oxide and nitrous oxide as electron acceptors in anaerobic respiration. The common denitrifying bacteria include several species of Pseudomonas, Alkaligenes and Bacillus. Their activities result in substantial losses of nitrogen into the atmosphere, roughly balancing the amount of nitrogen fixation that occurs each year. |
|
| Further information: websites International Symbiosis Society (not on this server) A valuable site that deals with the ecology and physiology of N fixation: http://web.reed.edu/academic/departments/biology/nitrogen/ (not on this server) A research-related site on Molecular genetics of plant-microbe interactions: http://www.lifesci.ucla.edu/mcdbio/html/ri4.htm (not on this server) |
This site is no longer maintained and has been left for archival purposes
Text and links may be out of date