This site is no longer maintained and has been left for archival purposes
Text and links may be out of date
Fungi are one of the three major groups
of eukaryotic organisms, equivalent in status to plants
and animals. They have major environmental and economic
significance:
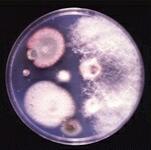
These and other aspects of fungal biology are discussed in various Profiles on this site. |
|
The fungal lifestyle We can think of fungi in a 'narrow' taxonomic sense or in terms of lifestyle. This distinction is necessary because some of the organisms that we regard as fungi - the extremely important potato blight pathogen (Phytophthora infestans) is one example - are quite unrelated to the true fungi and belong to a separate kingdom of organisms (see later). However, we can define the fungi broadly in terms of lifestyle and accommodate all these organisms.
|
|
|
Figure A. Tip growth of fungal hyphae. These three images from a videotape sequence were taken over a period of 20 minutes after a glass coverslip was placed on the margin of a colony of Neurospora crassa. The left-hand image is typical of a hypha, with extension growth at the hyphal apex, and branches arising from points behind the main apex. The addition of a coverslip disturbs the normal tip growth, causing several branches to emerge from the existing tips (centre and right-hand images). |
|
Apical growth is the hallmark of fungi (see Fungal tip growth and hyphal tropisms). It enables them to grow continuously into fresh zones of nutrients and also to penetrate hard surfaces such as plant cell walls, insect cuticle, etc. This is why fungi are so important as plant pathogens and as decomposer organisms. |
|
Figures B, C show two examples of potent fungal toxins (mycotoxins). B. A sheep suffering from facial eczema, a condition caused by the toxin sporidesmin in spores of the fungus Pithomyces chartarum. The fungus grows on pasture grasses in New Zealand, and sheep that ingest the spores develop liver damage leading to a photosensitive response (see Airborne microorganisms). [Image supplied by Dr Eric McKenzie]. C. Ergot of cereals, caused by the fungus Claviceps purpurea. This fungus infects cereals and grasses, forming large black resting bodies (ergots) in place of some of the grains. The ergots contain extremely toxic alkaloids that can lead to death of people who eat bread made from contaminated flour. Click here for further images and information on facial eczema. |
|
The major groups of fungi and fungus-like organisms A 'narrow' definition of fungi would include only the organisms in group 1 below. These are closely related, with a common ancestor, and represent the Kingdom Mycota. However, several other organisms have traditionally been treated as fungi. They are shown in groups 2 and 3 below. 1. True fungi (Kingdom Mycota) with walls typically containing chitin, and with many other characteristic cellular and biochemical features. Five sub-groups are recognised.
2. Organisms with a fungal lifestyle (Kingdom Stramenopila) but with cellulose walls and with cellular and biochemical features resembling those of plants. For further details, see Zoosporic fungi online (not on this server). The Kingdom Stramenopila contains brown algae, diatoms and some fungus-like organisms, including the Oomycota.
3. Organisms with some fungus-like features, but which grow as wall-less protoplasmic stages. There are several unrelated organisms in this group, loosely termed "slime moulds"
Click HERE (not on this server) for further details of fungal groups, including the fossil record and phylogenetic relationships |
|
|
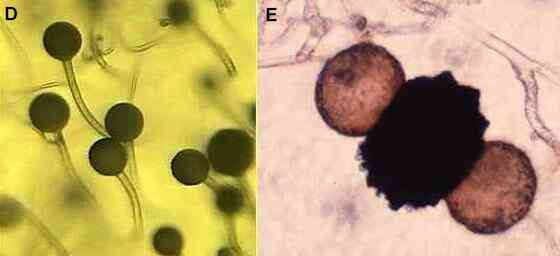
Figures D-E.
Zygomycota. Asexual
reproduction (D) is by production of spores in a sporangium
on an aerial hypha termed a sporangiophore
(see Fungal tip growth). The appearance of these
structures leads to the common name "pin
moulds". Sexual reproduction (E) is by fusion of two
gametangia to produce a black, warty zygospore,
with swellings on either side termed suspensors. |
|
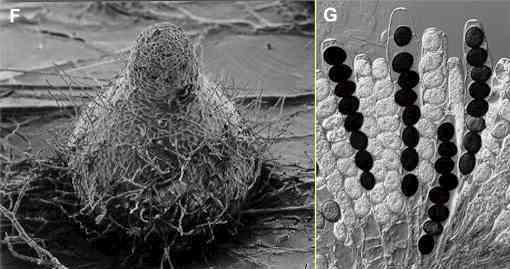
Figures F, G.
Ascomycota. Sexual
reproduction in this group leads to the production of one
or more asci, each containing usually 8 ascospores.
Figure G shows a cluster of asci of Sordaria
macrospora; the mature ascospores are black. Often
the asci are contained in a fruiting body which may be
flask-shaped (termed a perithecium and
shown for S. macrospora in Figure F)
or cup-shaped (an apothecium) or closed
(a cleistothecium). See Lichens for examples of apothecia; see Thermophilic
microorganisms for
an example of a cleistothecium. |
|
|
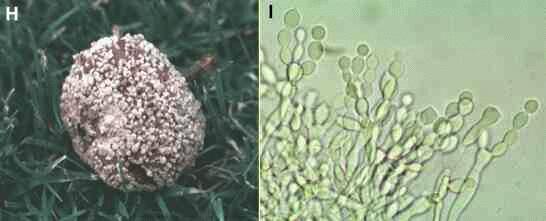
Figures H, I. Deuteromycota. Deuteromycota produce asexual spores (conidia) in many ways but never within a sporangium. Several examples can be found in Thermophilic microorganisms. The example here is Monilinia fructigena, a common cause of apple rots in orchards. It infects the fruit through wounds caused by birds or wasps, then rots the fruit rapidly and forms many sporing pustules on the surface (Figure H). At high magnification (Figure I) the spores are seen to develop in branched chains by successive budding and swelling of the hyphal tips. |
|
Figures J-N. Basidiomycota. The basidiomycota include the economically important rust fungi of crop plants and the mushroom- and toadstool-producers. Figure J shows toadstools of Coprinus comatus, one of the 'ink-caps'. In this and other Coprinus species the gills are digested progressively and drip down as an inky fluid containing the basidiospores (see Figure K, taken 2 days after Fig. J). Figures L and M show microscopical cross sections of the gills of a typical toadstool. The basidiospores (seen in Fig. M) are produced from basidia that line the gills. Figure N shows a different type of fruiting body - a puffball (Lycoperdon species) which contains many basidiospores. At maturity, the puffball dries to a 'papery' sack with a hole at the top, and the spores are puffed out by falling raindrops (see the book cover at the top of this page). |
|
|
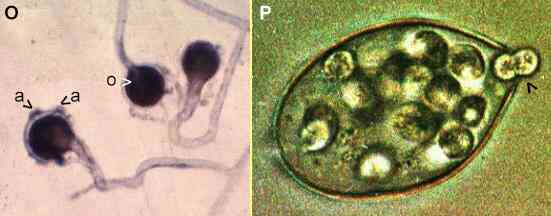
Figures O, P. Oomycota. Sexual reproduction is shown for a
species of Saprolegnia in Figure O.
An oogonium is fertilised by 'male'
hyphal branches that clasp onto it and produce antheridia
(a). Then the contents of the oogonium
are converted to one or more thick-walled resting spores
termed oospores (o).
Asexual reproduction is by release of motile zoospores
from a sporangium. Figure
P shows a single
sporangium of Phytophthora palmivora. This
fungus is related to P. infestans (the cause of
potato blight). The sporangia are wind-dispersed, land on
a plant surface and their contents differentiate to
produce zoospores. A small papillum at the tip of the
sporangium breaks down, and zoospores escape (arrowhead)
by swimming through the resulting pore. [Figure P
supplied by David Grayson] |
|
Distinctive features of the true fungi (Kingdom Mycota) The true fungi share several features that clearly distinguish them from all other organisms, showing that they are a 'natural' (monophyletic) group of organisms. These features are important in practice because they can provide targets for the actions of antifungal agents. In the list below we see that some of the most powerful drugs for treatment of fungal infections of humans, and the fungicides used for plant disease control, are targetted at the unique biochemical or structural features of fungi (see Penicillin and other antibiotics). 1. Chitin is a major component of fungal walls (but also found in insects, etc.). The enzyme that synthesizes chitin (chitin synthase) is a target for the polyoxin antibiotics. 2. Fungi are haploid, whereas the other major groups of eukaryotes are diploid. 3. Fungal cell membranes contain ergosterol, whereas animals have cholesterol and plants have sitosterol and other 'phytosterols'. Several antifungal drugs (e.g. ketoconazole) used in human therapy act by blocking ergosterol synthesis. The antifungal antibiotics (e.g. nystatin, amphotericin B) combine with ergosterol in fungal membranes. And several fungicides used for plant disease control act by disrupting specific steps in the ergosterol synthesis pathway. 4. Fungi synthesise the amino acid lysine by a unique pathway, different from that of other organisms. 5. Fungi have characteristic soluble carbohydrates (the disaccharide trehalose and polyhydric alcohols like mannitol and arabitol) and storage compounds (e.g. glycogen), differing from those of most plants and animals. 6. Fungi have several characteristic ultrastructural features, such as plate-like cristae in the mitochondria (like animals), and tubular unstacked Golgi cisternae (unlike animals or plants). See Fungal tip growth. 7. The microtubules of fungi have unique binding affinity for anti-tubulin agents. In particular, fungal tubulins bind to the antibiotic griseofulvin (used to treat some fungal infections of humans) and to the benzimidazole fungicides (used widely for control of fungal pathogens of plants). 8. Finally, fungi differ from other organisms in a range of biochemical and molecular features such as the regulation of some enzymes, some aspects of mitochondrial codon usage, etc. |
This site is no longer maintained and has been left for archival purposes
Text and links may be out of date