This site is no longer maintained and has been left for archival purposes
Text and links may be out of date
| 1. Orchid mycorrhizas Some types of orchid are non-photosynthetic; others only produce chlorophyll when they have grown past the seedling stage. In all cases, the plant depends on sugars derived from a fungal partner for at least part of its life. The minute orchid seeds, with negligible nutrient reserves, will not germinate unless a fungus infects them, although the seeds can germinate aseptically if supplied with the 'fungal sugar' trehalose. These mycorrhizas are unusual because, in effect, the plant parasitises the fungus that invades it. The fungi in these associations resemble the common plant pathogen Rhizoctonia solani, but recent taxonomic studies have assigned them to several related genera. They are mainly saprotrophic - they grow by degrading organic matter in soil - but they might obtain trace elements or some other factor from the plant. An interesting variation on this theme is shown by some orchids and other non-photosynthetic plants (Monotropa species) that have wood-rotting species of Armillaria as their mycorrhizal symbionts. In some of these cases the fungus can even be a pathogen of tree roots, so that the non-photosynthetic plant gains its nutrients by indirect parasitism of a living tree (see Armillaria). Further reading: Anderson TF & Rasmussen HN (1996) The mycorrhizal species of Rhizoctonia. pp. 379-390 In Rhizoctonia species: Taxonomy, Molecular Biology, Ecology, Pathology and Disease Control (B Sneh et al., eds) Kluwer Academic, Dordrecht. |
|
 |
|
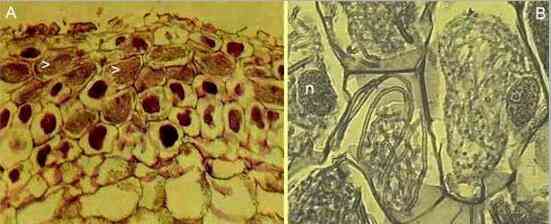
| Figure A (above) shows a cross-section of the outer part of the protocorm of an orchid, Neottia, stained to reveal the masses of fungal hyphae (arrowheads). Figure B shows part of a section at much higher magnification. The cells of the orchid are filled with coils of fungal hyphae but, significantly, the plant cells are still alive and they contain nuclei (n). The fungal coils will only last a few days or weeks before they are digested (those in the nucleate cell on the right of Figure B appear to be degenerate) and the process of invasion and digestion will begin again. | |
| 2. Arbuscular mycorrhizas Arbuscular mycorrhizas are found on the vast majority of wild and crop plants, with an important role in mineral nutrient uptake and sometimes in protecting against drought or pathogenic attack. Structures resembling those of the present-day AM fungi have been found in fossils of primitive pteridophytes of the Devonian period. It is thought that these fungi colonised the earliest land plants and that mycorrhizal associations could have been essential for development of the land flora. The fungi involved are
members of the zygomycota (related
to Mucor). They are classified currently in six
genera (Acaulospora, Entrophospora, Gigaspora,
Glomus, Sclerocystis and Scutellospora)
and they seem to be obligate symbionts: none of them can
be grown in axenic culture, i.e. in the
absence of their hosts. |
|
 |
|
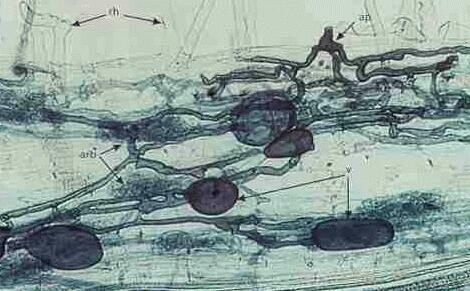
| The image above shows part of a clover root
from the Pentland Hills near Edinburgh, naturally
infected by an AM fungus. There was no evidence of fungal
infection until the root tissues were cleared with strong
alkali and then stained with trypan blue to reveal the
fungus. The site of penetration is shown at top right,
where the fungus produced a pre-penetration swelling (appressorium,
ap), then it grew between the root cells and formed
finely branched arbuscules (arb) and
swollen vesicles (v). The arbuscules are
thought to be sites of nutrient exchange - the fungus
obtains sugars from the plant, and the plant obtains
mineral nutrients (e.g. phosphorus) that the fungus
absorbs from the soil. Vesicles are thought to be used
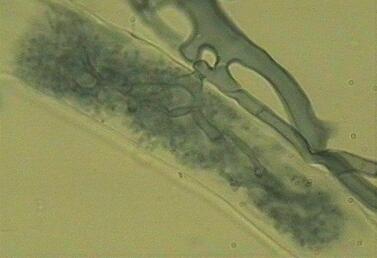
for storage. Root hairs (rh) are also labelled. The image below shows a single arbuscule with its repeated dichotomous branching inside a root cell. The plant cell remains alive, because its membrane extends to encase all the branches of the fungus. Strictly speaking, therefore, the fungus is always outside of the cell, surrounded by the cell membrane. Feeding relationships of this type, in which a fungus produces special nutrient-absorbing structures within the host cells, are termed biotrophic. For further details see Biotrophic plant pathogens. |
|
|
|
The hyphae of AM fungi extend into soil, where their large surface area and efficient absorption enable them to obtain mineral nutrients, even if these are in short supply or are relatively immobile. AM fungi seem to be particularly important for absorption of phosphorus, a poorly mobile element, and a proportion of the phosphate that they absorb has been shown to be passed to the plant. |
|
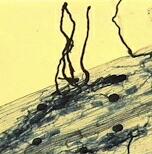
3. Ectomycorrhizas Ectomycorrhizas (sometimes termed ectotrophic mycorrhizas) are characteristic of many trees in the cooler parts of the world - for example pines, spruces, firs, oaks, birches in the Northern Hemisphere and eucalypts in Australia. However, some trees (e.g. willows) can have both ectomycorrhizas and arbuscular mycorrhizas, and most tropical trees have only arbuscular mycorrhizas. The fungi involved are
mainly Ascomycota and Basidiomycota, including many that produce the
characteristic toadstools of the forest floor (Figures
A-C below). Most of these fungi can be grown in
laboratory culture but, unlike the wood-rotting fungi,
they are poor degraders of cellulose and other plant wall
materials. So they gain most of their sugars from the
living plant roots in natural conditions. |
|
 |
|
A. Rings of toadstools of mycorrhizal fungi (Hebeloma and Lactarius species) around the base of a birch tree. B. Fruitbody of Lactarius. The gills of this toadstool were cut to show that the fungus exudes a milky fluid when the tissues are damaged - hence the name 'Lactarius'. C. Fruitbody of Amanita muscaria (the 'fly agaric') which is a typical mycorrhizal fungus of birches but also can be found on some other trees. D, E. Mycorrhizal roots. In ectomycorrhizas the terminal branches of the root system are highly modified - the roots are short and stumpy, covered with a mantle (sheath) of fungal tissue (the creamy-white root surface in D), and there are few or no root hairs. The fungus takes over the normal nutrient-absorbing role of the root hairs. In E the fungal mantle is less conspicuous, but the fuzzy appearance of the roots is due to many fungal hyphae growing from the mantle into the soil. Such roots are seen easily if the undecomposed, surface litter is scaped away from the forest floor to reveal the decomposing litter containing a mass of mycorrhizas and their fungal networks. F. Cross-section of a pine mycorrhiza, showing the substantial fungal sheath that encases the root (labelled 's').. G. Higher magnification of the sheath (left side) composed of a tightly packed fungal 'tissue'. From the inner side of the sheath, the fungus grows between the root cortical cells, forming a network termed the Hartig net (Hn). The sections in F and G were stained to show phenolic compounds (red) that often are formed in pine roots in response to mycorrhizal infection. They might have a role in limiting the fungal invasion of the tissues. |
|
Functioning of ectomycorrhizas Experiments with radioactive tracers have shown that when labelled CO2 is applied to leaves of tree seedlings, the label is found in plant sugars (sucrose, etc.) which move to the roots; then the label enters the fungal sheath where it occurs in the form of typical 'fungal carbohydrates' such as mannitol and trehalose. Most plants and plant tissues cannot metabolise these compounds, so there is, in effect, a one-way flow of carbohydrate to the fungus. The cost of this to the plant may be considerable. However, the plant also can benefit from the association, because the fungal hyphae that ramify into soil are very efficient in capturing mineral nutrients; these accumulate in the sheath but at least some minerals are transferred to the plant, presumably from the Hartig net. Recent work has shown that several ectomycorrhizal fungi can degrade proteins (they release protease enzymes) and thus can obtain nitrogen from the decomposing leaf litter. This could be highly significant in temperate and sub-boreal forests, where the rates of mineral nutrient recycling are low because of the low microbial activity in cool, acidic conditions. Ectomycorrhizal fungi might thus play a key role in the nitrogen nutrition of trees. It has also been shown that young seedlings growing in the shade of 'mother' trees can be attached to the 'parent' by a common network of mycorrhizal hyphae in the soil. In these conditions, at least some movement of labelled carbohydrates has been shown to occur from the roots of the parent to the younger trees, perhaps helping to nurture the seedlings. Further reading: MF Allen (ed) 1992. Mycorrhizal Functioning. Chapman & Hall, New York. Websites: Institute of Plant Nutrition, Hohenheim: http://www.uni-hohenheim.de/institute/pflanzenernaehrung/hhyphae.htm An ectomycorrhizal site: http://mycor.nancy.inra.fr/MycorWeb.html |
This site is no longer maintained and has been left for archival purposes
Text and links may be out of date