This site is no longer maintained and has been left for archival purposes
Text and links may be out of date
| Winogradsky column The prokaryotic bacteria and archaea exhibit an astonishing metabolic diversity, which far exceeds that of animals, plants, fungi and other higher organisms. The prokaryotes literally keep our biological world turning by recycling all the mineral elements necessary for life support. Two famous microbiologists pioneered the study of these processes: Sergius Winogradsky (1856-1953) and Martinus Willem Beijerinck (1851-1931). In contrast to the pure culture studies of other pioneer microbiologists such as Louis Pasteur and Robert Koch, these workers studied the relationships between different types of microorganisms in mixed communities. |
|
| A simple laboratory demonstration - the Winogradsky column - illustrates how different microorganisms perform their interdependent roles: the activities of one organism enable another to grow, and vice-versa. These columns are complete, self-contained recycling systems, driven only by energy from light! | |
The columns (Figure A) are easy to set up with a glass or perspex tube, about 30 cm tall and 5 cm diameter. Mud from the bottom of a lake or river is supplemented with cellulose (e.g. newspaper), sodium sulphate and calcium carbonate, then added to the lower one-third of the tube. The rest of the tube is filled with water from the lake or river, and the tube is capped and placed near a window with supplementary strip lights. All the organisms are present initially in low numbers, but when the tubes are incubated for 2 to 3 months the different types of microorganism proliferate and occupy distinct zones where the environmental conditions favour their specific activities. We shall return to the 'true' columns later, but first we will use an idealised column shown below, and list some of the main activities that occur. |
|
 |
|
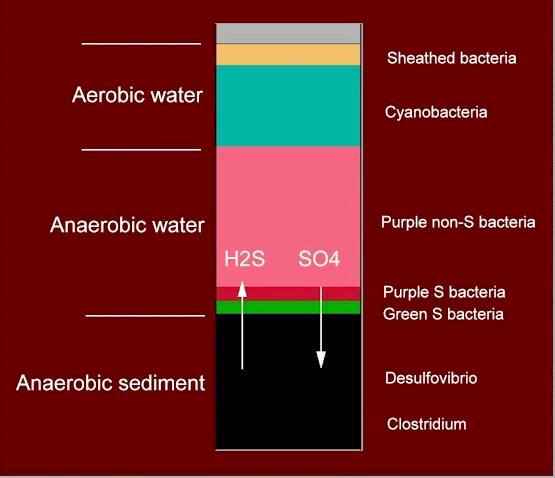
1. The large amount of cellulose added initially promotes rapid microbial growth which soon depletes the oxygen in the sediment and in the water column. Only the very top of the column remains aerated because oxygen diffuses very slowly through water. 2. The only organisms that can grow in anaerobic conditions are those that ferment organic matter and those that perform anaerobic respiration. Fermentation is a process in which organic compounds are degraded incompletely; for example, yeasts ferment sugars to alcohol. Anaerobic respiration is a process in which organic substrates are degraded completely to CO2, but using a substance other than oxygen as the terminal electron acceptor. Some bacteria respire by using nitrate or sulphate ions, in the same way as we use oxygen as the terminal electron acceptor during respiration. 3. Some cellulose-degrading Clostridium species start to grow when the oxygen is depleted in the sediment. All Clostridium species are strictly anaerobic because their vegetative cells are killed by exposure to oxygen, but they can survive as spores in aerobic conditions. They degrade the cellulose to glucose and then ferment the glucose to gain energy, producing a range of simple organic compounds (ethanol, acetic acid, succinic acid, etc.) as the fermentation end products. |
|
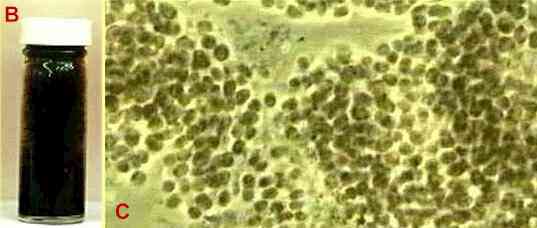
Figure B. This tube was filled with sterile nutrient medium containing sulphate, an organic acid and an iron nail. It was inoculated with a pure culture of Desulfovibrio, and after 10 days the contents turned black. Desulfovibrio has small, comma-shaped cells. Figure C. A species of Thiocapsa (purple sulphur bacterium) from the Winogradsky column (number 2) shown in Fig. A at the top of this page. Viewed by phase contrast microscopy. The purple colour can be seen where the cells are very dense. |
|
| 4. The sulphur-reducing bacteria (Fig. B) such as Desulfovibrio
can utilise these fermentation products by anaerobic
respiration, using either sulphate or other
partly oxidised forms of sulphur (e.g. thiosulphate) as
the terminal electron acceptor, generating large amounts
of H2S by this process.[In our own aerobic
respiration we use O2 and reduce it to H2O].
The H2S will react with any iron in the
sediment, producing black ferrous sulphide. This is why
lake sediments (and our household drains) are frequently
black. However, some of the H2S diffuses
upwards into the water column, where it is utilised by
other organisms. 5. The diffusion of H2S from the sediment into the water column enables anaerobic photosynthetic bacteria to grow. They are seen usually as two narrow, brightly coloured bands immediately above the sediment - a zone of green sulphur bacteria then a zone of purple sulphur bacteria (Fig. C). The green and purple sulphur bacteria gain energy from light reactions and produce their cellular materials from CO2 in much the same way as plants do. However, there is one essential difference: they do not generate oxygen during photosynthesis because they do not use water as the reductant; instead they use H2S. The following simplified equations show the parallel. 6 CO2 + 6 H20 = C6H12O6 + 6 O2 (plant photosynthesis) 6 CO2 + 6 H2S = C6H12O6 + 6 S (bacterial anaerobic photosynthesis) Actually, the balanced equation is: 6 CO2 + 12 H2S = C6H12O6 + 6 H20 + 12S The purple sulphur bacteria typically have large cells and they deposit sulphur granules inside the cells. The organism shown here (Fig. C) is a species of Thiocapsa. The green sulphur bacteria have smaller cells and typically deposit sulphur externally. The sulphur (or sulphate formed from it) produced by the photosynthetic bacteria returns to the sediment where it can be recycled by Desulfovibrio - part of the sulphur cycle in natural waters. |
|
6. Most of the water column above the photosynthetic bacteria is coloured bright red by a large population of purple non-sulphur bacteria. These include species of Rhodopseudomonas, Rhodospirillum and Rhodomicrobium. A mixed culture of them is shown in the bottle in Figure E (below). These bacteria grow in anaerobic conditions, gaining their energy from light reactions but using organic acids as their carbon source for cellular synthesis. So they are termed photoheterotrophs. The organic acids that they use are the fermentation products of other anaerobic bacteria (e.g. Clostridium species), but the purple non-sulphur bacteria are intolerant of high H2S concentrations, so they occur above the zone where the green and purple sulphur bacteria are found. The columns shown at the top of this page (Fig. A) have passed the stage where these organisms are common because the water columns became oxygenated by cyanobacteria (Fig. D). |
|
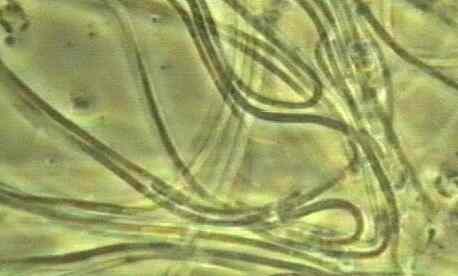
| Figure
D. Cyanobacterial filaments which
predominated in the upper region of column 2 (Fig. A)
|
|
7. Many microorganisms can grow in the oxygenated zone at the top of the water column, but three distinctive types are of special interest. (i) Any H2S that diffuses into the aerobic zone can be oxidised to sulphate by the sulphur-oxidising bacteria. These bacteria gain energy from oxidation of H2S, and they synthesize their own organic matter from CO2. So they are termed chemosynthetic organisms, or chemoautotrophs. Similar types of organism occur in soils, gaining energy from the oxidation of ammonium to nitrate, which then leaches from the soil and can accumulate in water supplies. (ii) Photosynthetic cyanobacteria can grow in the upper zones. These are the only bacteria that have oxygen-evolving photosynthesis like that of plants. In fact, there is very strong evidence that the chloroplasts of plants originated as cyanobacteria (or the ancestors of present-day cyanobacteria) that lived as symbionts inside the cells of a primitive eukaryote. Similarly, there is equally strong evidence that the mitochondria of present-day eukaryotes were derived from purple bacteria. Once the cyanobacteria start to grow they can oxygenate much of the water. This happened in column 2 (Fig. A) at the top of this page - the whole water column was dominated by a mass of cyanobacteria composed of spiral filaments (see Fig. D). |
|
(iii) The top of the water column can contain large populations of sheathed bacteria (se Column 1, Fig. A). These are aerobic organisms which use organic substrates, but are unusual because as the bacterial cells divide they synthesize a rigid tubular sheath from which individual cells can escape and swim away to establish new colonies. Many empty sheaths are seen in older colonies. They are made of a complex mixture of protein, polysaccharide and lipid, and are thought to protect the cells from predation by protozoa. The sheaths also can be encrusted with ferric hydroxide, giving a yellow or rusty appearance to the colonies. |
|
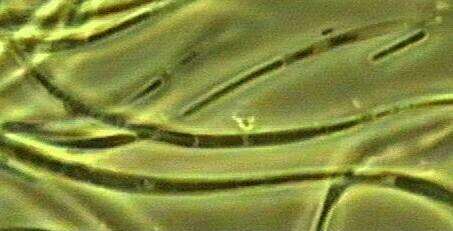
Narrow sheaths of the sheathed bacteria, taken from the upper yellow-orange zone of the Winogradsky column (labelled 1) in Fig. A. Most of the sheaths were empty. Same magnification as used for the cyanobacterium in Fig. D. |
|
Summary The Winogradsky column is a classic demonstration of the metabolic diversity of prokaryotes. All life on earth can be categorised in terms of the organism's carbon and energy source: energy can be obtained from light reactions (phototrophs) or from chemical oxidations (of organic or inorganic substances) (chemotrophs); the carbon for cellular synthesis can be obtained from CO2 (autotrophs) or from preformed organic compounds (heterotrophs). Combining these categories, we get the four basic life strategies: photoautotrophs (e.g. plants), chemoheterotrophs (e.g. animals, fungi), photoheterotrophs and chemoautotrophs. Only in the bacteria - and among the bacteria within a single Winogradsky column - do we find all four basic life strategies. The Winogradsky column is also a classic demonstration of how microorganisms occupy highly specific microsites according to their environmental tolerances and their carbon and energy requirements. And, finally, the column enables us to see how mineral elements are cycled in natural environments. We focused mainly on sulphur, but there are equivalent cycles for nitrogen, carbon and other elements. |
|
This site is no longer maintained and has been left for archival purposes
Text and links may be out of date