Control
of the Japanese beetle
In this
section we discuss the use of a bacterium, Bacillus
popilliae, to control a major introduced pest in the
USA.
Much of the
text below has been copied and updated from a book now
out of print [JW Deacon, 1983. Microbial Control of
Plant Pests and Diseases. Van Nostrand Reinhold,
Wokingham]
===============================================================
The Japanese beetle, Popillia
japonica (Figure
A), was accidentally
introduced into the USA early this century. Although it
is not a problem in its area of origin, the beetle causes
serious damage in the USA. It spread rapidly from the
initial sightings in New Jersey (1916) and today it is
found over roughly half of the country, in almost every
state east of the Mississippi. It is a problem as an
adult beetle because it feeds on a wide range of plants,
eating out the leaf tissues between the leaf veins (Figure B), and it accumulates on ripening fruit
causing substantial damage. It is also a problem in the
larval stage because the adult beetles lay their eggs in
grass turf and the grubs destroy the grass roots,
especially on new housing estates where natural enemies
are absent.
|
 Figure A. Adult
Japanese beetles, about 1-2 cm long. Figure B, feeding damage on foliage. Based on slides
provided by Fairfax Biological Laboratory.
|
By the 1930s the beetle problem had become so serious
that a search was begun for a control measure. This led
to the discovery of some naturally occurring diseased
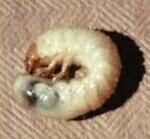
larvae. The disease was termed milky disease
because of the milky white appearance of the grubs, due
to a large number of refractile bacterial spores in the
haemolymph (insect blood) (Figures C, D). Two types of bacterium were subsequently
isolated from two types of milky disease. Type A disease
was characterized by a pure white appearance of the grubs
and the bacterium in this case was named B. popilliae.
Type B disease differed in that the grubs showed a
transition from white to brown over winter and the
bacterium causing this disease was named B.
lentimorbus. A range of other milky disease bacteria
were isolated from beetle hosts throughout the world, but
the trend now is to regard all of these as varieties of B.
popilliae because they are more closely
related to one another than they are to other Bacillus
spp.
All these
bacteria are specialised pathogens of beetles
(Coleoptera), specifically of the scarabaeid
beetles (family Scarabaeidae). This family includes the
beneficial dung beetles but also some of the most
important pasture pests - the chafers. In practice B.
popilliae has been used intensively and almost
exclusively for control of the Japanese beetle in the
USA, and to a lesser degree against the European corn
chafer Amphimallon majalis in that country.
The milky
disease bacteria are highly pathogenic and also highly
persistent in the environment so they can be used for
mass release to achieve lasting control. But B.
popilliae cannot be produced easily in artificial
media, so the inoculum for control programmes is produced
in living hosts.
|
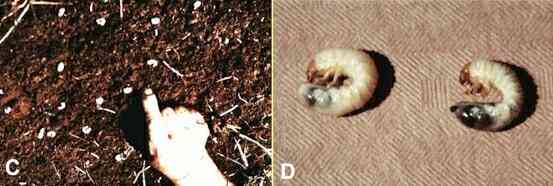 Figure C, larvae of the Japanese beetle in
soil; the grubs are about 2-3 cm long. Figure D, a healthy grub (right) and a diseased grub
(left). Based on slides provided by Fairfax
Biological Laboratory.
|
The
bacterium and its physiologyB. popilliae is a Gram-negative
spore-forming rod, 1.3 to 5.2 x 0.5 to 0.8 micrometres.
It is a fastidious organism that grows only on rich media
containing yeast extract, casein hydrolysate or an
equivalent amino acid source, and sugars. Several amino
acids are known to be required for growth, as are the
vitamins thiamine and barbituric acid. Trehalose, the
sugar found in insect haemolymph, is a favoured carbon
source though glucose also can be used.
Some varieties of B.
popilliae form a crystalline body inside the cell at
the time of sporulation and in this respect resemble B.
thuringiensis.
But the crystal is not thought to play a significant
role in infection and certainly it is not as important as
in B. thuringiensis. The variety lentimorbus, for
example, does not produce a crystal and yet it causes
disease. Another difference between B. popilliae and
B. thuringiensis is that B. popilliae cannot
be induced to sporulate in laboratory media although it
does so readily in the diseased host. Actually there are
a number of oligosporogenic mutants -
ones that produce a few spores - but spores for microbial
control programmes are usually produced in living insect
larvae - an expensive and time-consuming process.
|
The
host-parasite interactionB. popilliae causes disease of beetle
larvae when they ingest spores in the soil. The spores
germinate in the gut within 2 days and the vegetative
cells proliferate, attaining maximum numbers within 3 to
5 days. By this time some of the cells have penetrated
the gut wall and begun to grow in the haemolymph, where
large numbers of cells develop by day 5 to 10. A few
spores also are formed at this stage but in the variety popilliae
the main phase of sporulation occurs later and is
completed by 14 to 21 days when the larva develops the
typical milky appearance. In laboratory conditions the
larva remains alive until this stage and usually contains
about 5 x 109 spores. In field conditions,
however, there are reports that larvae sometimes die
earlier, before the main phase of sporulation is
completed. This is of concern because sporulation stops
when the host dies and the larva ultimately releases
fewer spores to maintain the level of infestation of a
site.
|
The cause of insect death is not fully known.
Physiological starvation caused by the growth of
bacterial cells in the haemolymph seems the most likely
explanation, and fat reserves of diseased larvae have
been shown to be much reduced compared with those of
healthy larvae. However, toxins also may be involved
because they have been detected in culture filtrates of
the bacteria and shown to be lethal on injection.
Recently, a crystal protein from sporulating cells of B.
popilliae was found to have similarities to one of
the Cry toxins of B. thuringiensis
(see B. thuringiensis). Although it does not cause such
drastic effects on the insect gut wall as do the B.
thuringiensis toxins, it might contribute to
pathogenic invasion through the gut wall (Zhang et al.,
1997).
|

Figures E-G. Injection
of healthy larvae of the Japanese beetle, as the first
stage in production of commercial spore powders. Based on
slides provided by Fairfax Biological Laboratory.
|
Application
for biological control B. popilliae has been registered for
control of the Japanese beetle in the USA since about
1950 - the first registration of any insect pathogen as a
microbial control agent. The control strategy is aimed
solely against the larvae, so if the beetle itself is
causing serious damage a chemical insecticide must be
used for short-term control. The bacterial spores are
produced commercially in larvae collected from grass turf
on golf-courses, airports, etc. The larvae are injected
with bacterial cells (Figures
E-G), incubated until
they develop a milky appearance and then crushed and
dried to give a spore powder (Figure H).
The spore powders are applied to turf in small heaps at
roughly 1-metre spacing (Figures I, J) and the spores are then distributed
naturally by wind and rain. They can persist in soil for
several years and infect larvae that eat them. Therefore
they have the potential to give lasting control of a pest
problem, because the spore numbers in soil are boosted
periodically when a diseased larva dies.
Commercial "milky
spore" powders are marketed under several names, by
several companies. For example, Fairfax Biologicals
markets its product under the trade name "Doom".
Other products include "Milky Spore",
"Grub Attack" and "Grub
Killer".
|

Figure H. After the larvae have been injected
with B. popilliae and incubated to develop milky
disease, they are crushed, flash-dried and mixed with a
diluent, to produce a commercial spore powder. This
powder is applied to the surface of turf (Figures I, J) where it will be washed into the
ground. Based on slides provided by Fairfax Biological
Laboratory.
|
The use of B.
popilliae has proved remarkably successful. Between
1939 and 1953 over 100 tons of spore powder were applied
to turf in over 160,000 sites in the USA as part of a
Government programme (Fleming, 1968). Larval numbers in
the turf were reduced 10- to 20-fold and the population
stabilized at this new low level, with corresponding
reductions in the levels of adult beetle damage. However,
the treatment is most effective when applied on a region-
or state-wide basis (or at least to relatively large
areas) to reduce overall the levels of beetle
infestation. It is less appropriate for use by small
landowners, who may control the larvae in their own turf
only to find their trees and shrubs being eaten by
beetles from their neighbours' properties. Also, because B.
popilliae is obligately dependent on its hosts for
sporulation and because some larvae may not ingest spores
(or not ingest enough to cause disease) a periodic
resurgence and decline of the pest problem can be
expected. The success of the control programme must be
judged not on this basis but by the fact that over a
number of years the mean level of pest damage is lower
than it would be in the absence of B. popilliae.
|
Advantages
and disadvantages of B. popilliaeThe advantages of B. popilliae
include (1) its very narrow host range (which is
environmentally desirable) and its consequent lack of
effect on beneficial insects; (2) its complete safety for
man and other vertebrates (for example, it does not grow
at 37oC); (3) its compatibility with other
control agents including chemical insecticides and, more
recently, insect-pathogenic nematodes (Thurston et al.,
1994); (4) its persistence, giving lasting control.
Its disadvantages,
however, include (1) the high cost of production in
vivo; (2) its slow rate of action; (3) most
importantly, its lack of effect on adult beetles which
often cause the most obvious and distressing damage, and
(4) its relative unattractiveness to the small landowner.
Outstanding
problems
There is evidence that the
Japanese beetle has re-emerged as a serious pest in some
regions where it had been controlled effectively since
the initial applications of spore dust in the 1940s
(Dunbar & Beard, 1975). Larval densities ranged from
0 to 474 per square metre of turf in 1974 (mean 112), and
were sometimes as high as those recorded 25 years
earlier, before the control programme was begun.
Moreover, in this study only 0.2% of larvae collected
from field sites showed symptoms of milky disease
compared with 41.5% disease incidence in a survey in 1946
after B. popilliae had been introduced. Spores
collected from these few diseased larvae caused only 7 to
17% infection of larvae in laboratory tests, compared
with 65 to 67% infection from spores collected from New
York State where a decline in the degree of control had
not been reported. Even this figure was low in relation
to the expected 90% disease incidence at the inoculum
level used. Perhaps there has been a reduction in
virulence of B. popilliae in field sites over the
years, coupled with an increased degree of resistance of
the target pest (see Redmond & Potter, 1995). This
might be expected by natural selection, because an
obligately pathogenic bacterium that kills its host too
rapidly would be at a selective disadvantage.
|
Further
readingArticles:
WE Fleming (1968)
Biological Control of the Japanese Beetle. United
States Department of Agriculture Technical Bulletin
Number 1383. Washington DC.
DM Dunbar & RL
Beard (1975) Present status of milky disease of
Japanese and other Oriental beetles in Connecticut. Journal
of Economic Entomology 68,
453-457.
GS Thurston, HK Kaya
& R Gaugler (1994) Characterizing the enhanced
susceptibility of milky-disease infected scarabaeid
grubs to entomopathogenic nematodes. Biological
Control 4, 67-73.
CT Redmond & DA
Potter (1995) Lack of efficacy of in-vivo and
putatively in-vitro produced Bacillus popilliae
against field populations of Japanese bettle
(Coleoptera: Scarabaeidae) grubs in Kentucky. Journal
of Economic Entomology 88,
846-854.
JB Zhang, TC Hodgman,
L Krieger, W Schnetter & HU Schairer (1997)
Cloning and analysis of the first cry gene from Bacillus
popilliae. Journal of Bacteriology 179,
4336-4341.
Websites
There are many websites on
milky disease (type "Japanese+beetle" or
"milky+spore" in a search engine). Here are
just a few:
Managing the Japanese beetle (not on this server)
Ornamentals and Turf: Insect Pest
Management (not on this server)
Milky Spore Japanese Beetle Control -a commercial supplier's site (not on this server)
GO
TO FULL LIST OF PROFILES?
|