Introduction The fungal group basidiomycota is best known for the production of large fruitbodies such as the mushrooms, puffballs, brackets, etc. However, the group also contains some microscopic fungi, including the important rust fungi and smut fungi that parasitise plants (see Biotrophic parasites), and some yeasts. One of these yeasts, Sporobolomyces roseus, is very common on moribund leaf surfaces and is a cause for concern because its basidiospores are respiratory allergens. Another is Cryptococcus neoformans, which grows commonly on old "weathered" bird droppings, and which can cause a fatal systemic infection of immunocompromised people. Its air-borne basidiospores initiate infection via the lungs, leading to the disease termed cryptococcosis. This is now the fourth most important life-threatening diseases of AIDS patients in the USA, and common also in Europe. Many of the basidiomycota with the larger fruitbodies (toadstools etc.) are common and important agents of wood decay (see Wood decay) or decomposers of leaf litter, animal dung, etc. Others are mycorrhizal on forest trees (see Mycorrhizas). Several species are grown commercially for food, including the common cultivated mushroom, Agaricus bisporus, and some more exotic species which can be found on supermarket shelves. For example, the supermarket package below contains the brown-coloured Shiitake mushroom (Lentinus edodes, traditionally grown in south-east Asia) and a mixture of grey, pink and yellow forms of the oyster fungus (Pleurotus ostreatus, reportedly an aphrodisiac).
|
||||
| Distinctive features and life cycle
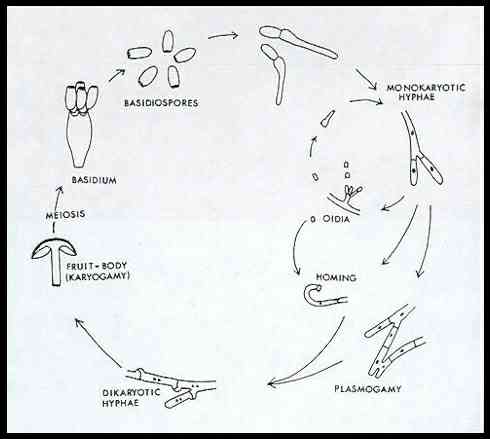
of the basidiomycota As a group, the basidiomycota have some highly characteristic features, which separate them from other fungi. They are the most evolutionarily advanced fungi, and even their hyphae have a dinstinctly "cellular" composition. This point is best illustrated by the life cycle below.
The basidiospores usually have a single haploid nucleus. When these spores germinate they produce hyphae with usually a single nucleus in each compartment. Because these hyphae have only one nuclear type they are termed monokaryons (from the Greek, mono = one; karyos = kernel or nucleus). At some stage in their growth, two monokarons of different compatibility groups (mating types) fuse. This can occur either by simple hyphal fusions or by fusion of a hypha with a small spore termed an oidium. The fusion event is termed plasmogamy. Then the nuclei divide and the daughter nuclei pair so that each hyphal compatment comes to contain two nuclei - one of each mating type. At this stage the fungus is termed a dikaryon (i.e. with two nuclear types). Many basidiomycota grow for most of their lives as dikaryons, until environmental signals induce them to produce fruibodies, such as a toadstool. All the hyphae that make up the toadstool are dikaryotic. At a late stage of development, some of these hyphae produce special cells termed basidia (singular, basidium). For example, the cells that line the gills of the common mushroom are basidia. Finally, the two haploid nuclei in each basidium fuse - a process termed karyogamy) to form a diploid nucleus. This then undergoes meiosis to produce four haploid nuclei, and these haploid nuclei migrate into the basidiospores, which develop on small stalks (termed sterigmata) from each basidium (see the image below). The dispersal of these monokaryotic spores completes the life cycle.
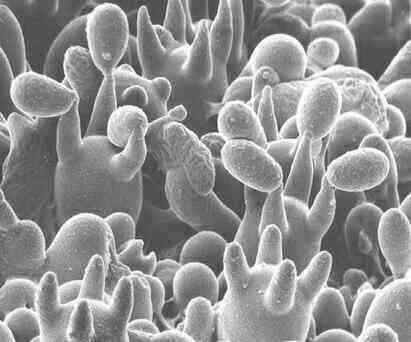
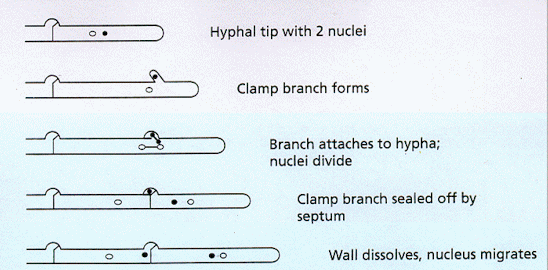
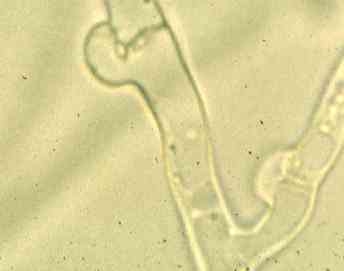
Scanning electron micrograph of basidia on the gills of a toadstool. Note that each basidium produces four stalks (sterigmata) and the basidiospores develop on the ends of these stalks. [Image courtesy of Dr C. Jeffree] In many basidiomycota there is a rather elaborate mechanism for ensuring that the dikaryotic condition is maintained during growth of the hyphae. As shown in the diagram below, the two nuclei in each tip cell divide at the same time, but one divides along the axis of the hypha, and the other divides so that a daughter nucleus enters a small, backwards-projecting branch. Then a septum forms to separate the original apical cell into two cells, and the branch fuses with the sub-terminal cell so that the nucleus in the branch migrates into this cell. The small branches at each septum are termed clamp connections. They can be seen at high magnification of a normal compound microscope (see the image below the diagram).
|
||||
Some unusual fruitbodies Many basidiomycota produce a typical toadstool-shaped fruitbody, with basidia lining the gills (or sometimes pores instead of gills). But an assemblage of basidiomycota termed gasteromycetes produce some unusual types of fruitbody, with basidia that mature inside the fruitbody. Puffballs (see below) are common examples of this - the basidiospores form a powdery mass inside the fruitbody. At maturity, the outer layer of the puffball becomes a papery structure with a hole at the top, and a "puff" of basidiospores is released when raindrops impact onto the fruibody.
Immature puffballs developing on rotten wood. Note the thick white strands on the surface of the wood. Each of these "mycelial strands" is composed of many hyphae, which channel nutrients to the developing fruitbodies from the mycelium growing in the wood.
Earthstars (above and below) are the fruitbodies of Geastrum and similar genera. They are like puffballs, but often develop from mycelium growing below ground in woodlands. In order to ensure that the fruitbodies are not covered in leaf litter, the outer layers of the fruitbody split and fold back at maturity, raising the inner "puffball" above ground level.
The "bird's nest fungi (see below) commonly grow on wood chippings. Their fruitbodies consist of a splash-cup, about 1 cm diameter. Within this are clusters of "eggs", termed peridioles. These are forcibly ejected when raindrops fall into the splash-cup, and they land on the surrounding ssubstrate, where they adhere by means of a sticky pad (termed a hapteron).
|
||||
| Fairy ring fungi Several basidiomycota grow as decomposers on the "thatch" of dead leaves and roots that accumulate in old grasslands. Their produce rings of fruitbodies in autumn, known as fairy rings because folklore associates them with the circles of fairy dances. The rings expand each year, as the fungal hyphae grow into new zones of thatch, and the hyphae in the older zone die, releasing nitrogen into the soil. This nitrogen promotes the growth of grass behind the advancing margin of the ring, giving it a richer green colour. Thus, we can see fairy rings for much of the year, as rings of enhanced grass growth, even when the fungal fruitbodies are not present. Some fairy rings are extremely large - for example those on the grass plains around Stonehenge in Wiltshire - and measurements of their current annual rates of expansion suggest that such rings can be at least 400 years old. Most fairy ring fungi produce only "green" rings and cause no damage to the grass. For example, this is true of the rings caused by Hygrocybe species, the fruitbodies of which are waxy and often brightly coloured.
However, the fungus Marasmius oreades (known as the fairy ring champignon; see below) forms such a thick, water-repellent mass of hyphae beneath the soil surface that it prevents water from penetrating the soil and therefore leads to a ring of dead turf, with a ring of darker, lusher grass behind this. M. oreades is known to produce hydrogen cyanide in laboratory culture, and HCN also has been detected in the killing zone of rings. But there is uncertainty about whether this contributes to death of the turf. In fact, the simplest way of overcoming the killing action of this fungus is to water the affected zone of turf with a dilute solution of a surfactant such as washing-up liquid.
|
||||