..
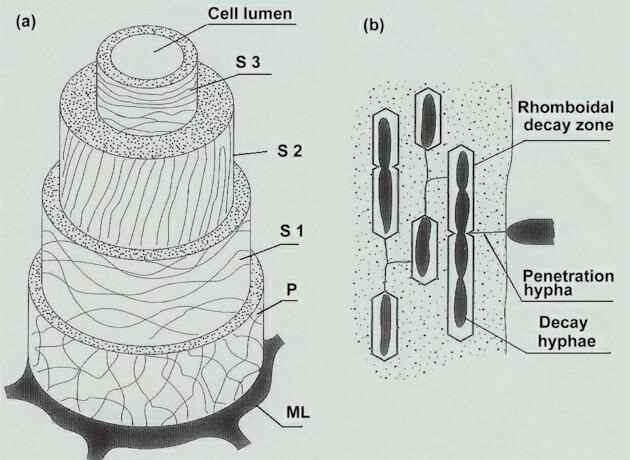
WOOD DECAY, AND WOOD-ROTTING FUNGI In terms of both its physical and chemical properties, wood is an exceptionally difficult substrate to degrade. One of the principal reasons is that wood contains very low levels of nitrogen, which is needed to produce the enzymes that degrade the main structural polmers of wood - cellulose (about 40-50% of the dry weight of wood), hemicelluloses (25-40%) and lignin (20-35%). The lignin component also presents a barrier to wood decay because lignin is a complex aromatic polymer that encrusts the cell walls, preventing access of enzymes to the more easily degradable cellulose and hemicelluloses. In addition to these points, wood often contains potentially fungitoxic compounds, which are deposited in the heartwood. In broad-leaved trees the toxic compounds are usually tannins, well know for their ability to cross-link proteins, making animal skins resistant to decay. In contrast, conifers contain a range of phenolic compounds such as terpenes, stilbenes, flavonoids and tropolones. The most toxic of the tropolones are the thujaplicins which act as uncouplers of oxidative phosphorylation; they are particularly abundant in cedarwood, making this a naturally decay-resistant wood for high-quality garden furnishings, etc. Despite this formidable list of obstacles, woody tissues are degraded by fungi, and these wood-decay fungi fall into three types according to their mode of attack on the woody cell walls - soft-rot fungi, brown-rot fungi and white-rot fungi. Soft-rot fungi Soft-rot fungi grow on wood in damp environments. They are the characteristic decay fungi of fence posts, telegraph poles, wooden window frames, the timbers of cooling towers, and wood in estuarine or marine environments. They have a relatively simple mode of attack on wood, illustrated in Fig. 1. Their hyphae grow in the lumen of individual woody cells, usually after entering through a ‘pit’ (depression) in the wall. Then they produce fine penetration branches that grow through the thin, lignin-coated S3 layer of the wall, to gain access to the thick, cellulose-rich S2 layer. When the penetration hyphae find a longitudinal plane of weakness in the S2 layer, they produce broader T-shaped hyphae which grow along the plane of weakness and secrete cellulase enzymes. The diffusion of these enzymes creates a characteristic pattern of decay, seen as rhomboidal cavities within the cell wall. These persist even when the fungi have died, leaving the characteristic ‘signature’ of a soft-rot fungus. The soft-rot fungi have little or no effect on lignin, which remains more or less intact. All the soft-rot fungi need relatively high nitrogen levels for wood decay, typically about 1% nitrogen content in the wood. If this is unavailable in the wood itself, then nitrogen can be recruited from the environment, such as the soil at the bases of fence posts, etc. The fungi that cause soft rots include several Ascomycota and mitosporic species, such as Chaetomium and Ceratocystis in terrestrial environments and species of Lulworthia, Halosphaeria and Pleospora in marine and estuarine environments.
Fig 1. (a) Diagram of the cell wall layers in woody tissue, showing the arrangement of cellulose microfibrils. ML = middle lamella between adjacent woody cells; P = thin primary wall with loosely and irregularly arranged microfibrils; S1-S3 = secondary wall layers. (b) Characteristic decay pattern of a soft-rot fungus in the S2 layer. The fungus penetrates by narrow hyphae, then forms broader hyphae in planes of weakness in the wall, and these hyphae produce rhomboidal cavities where the cellulose has been enzymatically degraded. [© Jim Deacon] Brown-rot fungi Brown-rot fungi are predominantly members of the Basidiomycota, including common species such as Schizophyllum commune, Fomes fomentarius (the ‘hoof fungus’ of Scottish birch woods; see Chapter XX) and the ‘dry-rot fungus’, Serpula lacrymans (Chapter 7). Many of the brown-rot fungi produce bracket-shaped fruitbodies on the trunks of dead trees, but the characteristic feature of these fungi is that the decaying wood is brown and shows brick-like cracking – a result of the uneven pattern of decay, causing the wood to split along lines of weakness (See Fig. 2). The term ‘brown rot’ refers to the characteristic colour of the decayed wood, because most of the cellulose and hemicelluloses are degraded, leaving the lignin more or less intact as a brown, chemically modified framework.
Fig 2. Part of a pine stump showing the characteristic brick-like decay by brown-rot fungi. [© Jim Deacon] The hyphae of brown-rot fungi occur very sparsely in the wood, often restricted to the lumen of woody cells, and yet they cause a generalised decay in which the S2 wall layer is almost completely degraded. This type of decay cannot be explained by the diffusion of cellulase enzymes, which are too large to diffuse very far, and too large even to pass through the pores in the S3 layer. In fact, the cellulases of brown-rot fungi have little effect on cellulose in vitro, unlike the cellulases of soft-rot fungi. Instead, the brown-rot fungi degrade cellulose by an oxidative process, involving the production of hydrogen peroxide during the breakdown of hemicelluloses. Being a small molecule, H2O2 can diffuse through the woody cell walls to cause a generalised decay. In support of this, the characteristic decay pattern of brown-rot fungi can be mimicked experimentally by treating wood with H2O2 alone, and at least one of these fungi, Poria placenta, has been shown to degrade cellulose only if hemicelluloses also are present, as substrates for generating H2O2. This mode of attack is an efficient way of using the scarce nitrogen resources in wood, because it does not require the release of large amounts of extracellular enzymes. White-rot fungi White-rot fungi are more numerous than brown-rot fungi. They include both Ascomycota, such as Xylaria spp.(Fig. 3), and Basidiomycota (e.g. Armillariella mellea).
Fig 3. Upper row: Two common Ascomycota that cause white rots. Left: Xylaria hypoxylon, the ‘candle snuff’ fungus often seen on rotting stumps. The upper parts of the fork-shaped structures are covered with white, powdery conidia. Right: Xylaria polymorpha (‘dead man’s fingers’) which often grows from the bases of rotting wood stumps. Bottom: A section cut through one of the “dead man’s fingers”, showing numerous perithecia just below the surface of the fruitbody.
Fig 4. Small, leathery, bracket-shaped fruitbodies of the white-rot fungus Coriolus versicolor, growing in an unexpected setting – is nothing sacred!? The white-rot fungi seem to use conventional cellulase enzymes for wood decay, but they are extremely efficient in their use of nitrogen. For example, the nitrogen content of Coriolus versicolor is about 4% when the fungus is grown on laboratory media of Carbon-to-nitrogen ratio, 32:1, but only 0.2% when grown on a medium of C:N, 1600:1. In nitrogen-poor conditions this fungus seems preferentially to allocate nitrogen to the production of extracellular enzymes and essential cell components, and it also efficiently recycles the nitrogen in its mycelia. White-rot fungi might also benefit from the growth of nitrogen-fixing bacteria in wood. The most remarkable feature of white-rot fungi is their ability completely to degrade lignin – they are the only organisms known to do this. As shown in Fig. 11.22, lignin is a complex polymer composed of three types of phenyl-propane unit (six-carbon rings with three-carbon side chains) bonded to one another in at least 12 different ways. If lignin were to be degraded by conventional means it would require a multitude of enzymes. Instead, lignin is degraded by an oxidative process. The details of this are complex, but essentially the white-rot fungi produce only a few enzymes (lignin peroxidase, manganese peroxidase, H2O2-generating enzymes, and laccase) and these generate strong oxidants, which virtually “combust” the lignin framework (Kirk & Farrell, 1987). The major enzyme that initiates ring-cleavage is laccase, which catalyses the addition of a second hydroxyl group to phenolic compounds. The ring can then be opened between two adjacent carbon atoms that bear the hydroxyl groups (Fig. 11.23). This process occurs while the ring is still attached to the lignin molecule. It is termed ortho fission, in contrast to meta fission which bacteria employ to cleave the phenolic rings of pesticide molecules (where the ring is opened at a different position - see Fig. 11.23). The other enzymes are involved mainly in generating or transferring oxidants. They include glucose oxidase which generates H2O2 from glucose, manganese peroxidase which oxidises Mn (II) to Mn (III), and which can then oxidise organic molecules, and lignin peroxidase which catalyses the transfer of singlet oxygen from H2O2 to aromatic rings and is one of the main initiators of attack on the lignin framework. These initial oxidations involving single electron transfers generate highly unstable conditions, setting off a chain of chemical oxidations. In addition to the fungi mentioned above, several others are commonly found on stumps or on the decaying major roots of trees. Examples of these include the distinctive Pholiota squarrosa, the very common "sulphur tuft, Hypholoma fasciculare and the "Velvet shank", Flammulina velutipes. GO TO MORE WOOD-ROTTING FUNGI ? (Pholiota, Hypholoma, Flammulina) |