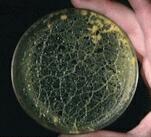
Until recently, the slime moulds were regarded as organisms of uncertain taxonomic status, claimed as fungi by mycologists and as protozoa by protozoologists. To make things worse, several groups of organisms have been included in the general term 'slime mould'. They include the cellular slime moulds which are unicellular amoeboid organisms such as Dictyostelium; the endoparasitic slime moulds such as the damaging plant pathogen Plasmodiophora brassicae (clubroot disease of cruciferous crops), and the plasmodial slime moulds such as Physarum species. Analysis of DNA sequences has now shown clearly that all these groups are evolutionarily distinct from one another, and also distinct from the fungi. So they can be regarded as separate kingdoms of organisms or separate lineages within the protozoan group (See "Tree of life"). Here we consider just one of these groups - the plasmodial slime moulds. We will call them by one of their formal names - the myxomycota. The myxomycota grow as a naked network of protoplasm, engulfing bacteria or other small food particles by phagocytosis. They are common in moist, organic-rich environments and especially on damp, rotten wood, where there is an abundance of bacteria as a food source. However, they are seldom seen in nature until they begin to sporulate, when they can produce conspicuous and brightly coloured fruiting bodies. These ripen to release masses of small, walled spores resembling fungal spores. For this reason, the slime moulds were once considered to be fungi. |
|
 |
|
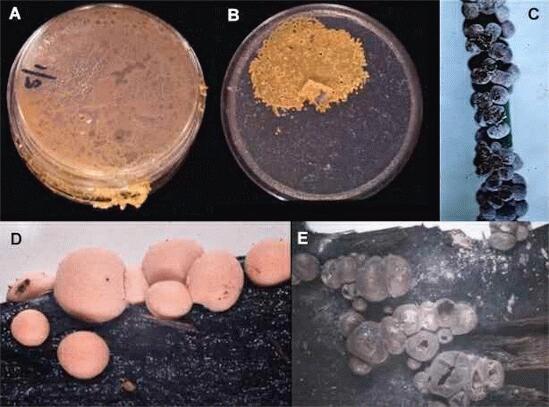
Figure A shows a colony of Physarum polycephalum, one of the few slime moulds that can be grown easily on agar media. The colony consists of a multinucleate network of protoplasm, termed the plasmodium. There is no wall, only a plasma membrane and a gelatinous sheath around this. Once the plasmodium has grown to fill the agar plate, it will migrate and start to move out of the dish or (B) accumulate all the protoplasm in a localised zone, leaving only the gelatinous sheath over the rest of the agar surface. The sheath is known to contain actin, which might be involved in migration. Under a microscope, the tubes (veins) that make up the plasmodium are seen to have gel-like protoplasm towards the outside, and rapidly streaming sol-like protoplasm in the centre. The rate of protoplasmic streaming can be extremely fast - over 1 mm per second - and it is rhythmic because its direction reverses every few seconds. At the onset of starvation, and in response to other environmental triggers, the plasmodium migrates to an aerial environment and converts to one or more fruiting structures termed sporangia (C-E). These usually are brightly coloured. Figure C shows the small fruiting structures of Physarum cinereum on a grass leaf. This species is quite commonly seen as grey patches on lawns in damp conditions, but it uses the grass blades merely as a 'perch' on which to fruit, and it can simply be brushed off the lawn. Figures D and E show two stages in the fruiting of Lycogala epidendrum, a common species on rotten wood. D is the early stage of development in which the fruiting bodies are bright pink; at this stage they feel spongy and they release a pink fluid, which in some way must have given this species its common name in the USA - 'Wolf's-milk fungus'. E is the final stage, where the sporangia have matured and turned grey. They release small grey-coloured spores by rupture of the sporangial coat. After being wind-dispersed, the walled spores germinate to release either an amoeboid cell - the myxamoeba - or a flagellate swarmer cell. These two cell types can interconvert. The myxamoebae can multiply to large numbers, but in many slime moulds they will not produce plasmodia unless two myxamoebae of different mating types fuse with one another. Thus, the myxamoebae seem to be haploid, and the plasmodial stage is diploid. |
|
| The myxomycota are not specially important or notable for their activities in natural environments, but their peculiar behaviour has made them important as model systems for experimental studies on topics such as the cell cycle, morphogenesis and chemotaxis. | |
Further reading: WF Dove, J Dee, S Hatano, FB Haugli & K-E Wohlfarth-Botterman (1986) The Molecular Biology of Physarum polycephalum. Plenum, New York. JP Armitage & JM Lackie (eds) (1991) Biology of the Chemotactic Response. Society for General Microbiology Symposium 46. Cambridge University Press, Cambridge. MF Madelin (1990) Methods for studying the ecology and population dynamics of soil myxomycetes. Methods in Microbiology 22, 405-416. |