..
IMAGES FROM CHAPTER 7: FUNGAL METABOLISM AND FUNGAL PRODUCTS
Fig. 7.1. Overview of the central metabolic pathways of fungi, showing how the main energy-yielding pathway (the Embden-Meyerhof pathway and the tricarboxylic acid cycle) provide the precursors for biosynthesis of various metabolic products (shaded boxes). Note that only some of the intermediates of the central metabolic pathway are shown - see Fig. 7.2 for more details. Also, note that secondary metabolites (including penicillins and mycotoxins) are produced from various precursors, but primarily from acetyl coenzyme A. [© Jim Deacon]
Fig. 7.2. Outline of the Embden-Meyerhof pathway and tricarboxylic acid cycle, which provide the major means of generating energy from sugars. Also shown (top left) is the pentose phosphate pathway which can provide some energy, but its major role is for biosynthesis, including synthesis of the 5-carbon sugars of nucleic acids. [© Jim Deacon]
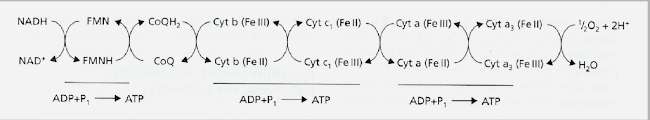
Fig. 7.3. Outline of the respiratory electron transport chain (CoQ = coenzyme Q, Cyt = cytochrome). The energy yield from aerobic respiration We can calculate the theoretical energy yield from glucose during aerobic respiration by calculating the number of ATP molecules that could be synthesised (see Fig. 7.2) as follows::
However, the actual ATP yield will be much less than this, for at least two reasons.
We should mention the special role of the pentose-phosphate pathway (Fig. 7.2). It can be used as an alternative to the EM pathway for generating energy from sugars (giving 1 ATP instead of the 2 ATP from the EM route). But its major role is in biosynthesis - it generates some important intermediates, such as ribose-5-phosphate for the synthesis of nucleic acids and erythrose-4-phosphate for the synthesis of aromatic amino acids. What happens when oxygen is limiting? In the absence of a terminal electron acceptor (usually oxygen), the electron transport chain cannot operate, so the pool of reduced nucleotides (NADH, etc.) accumulates and metabolism would rapidly cease. But fungi, and many other organisms, can still obtain some energy in the absence of oxygen by one of the following reactions.
In both cases the end-product (ethanol or lactic acid) is more reduced than pyruvic acid, so the reactions are coupled with the reoxidation of NADH to NAD+. This allows the Embden-Myerhof pathway to continue, and the cells release either lactic acid or ethanol into the surrounding medium.Most yeasts and mycelial fungi produce ethanol – this is the basis of the alcoholic drinks industry. But several Chytridiomycota produce lactic acid (e.g. Allomyces, Blastocladiella), as do humans when our tissue oxygen level is depleted. Energy-yielding reactions of this type – where an internal inorganic compound is the terminal electron acceptor – are defined by the biochemical term, fermentation. In equation 1 (above) the terminal electron acceptor is acetaldehyde, because this accepts electrons from NADH and is, itself, reduced to ethanol. Similarly, in equation 2, pyruvic acid accepts electrons and is itself reduced to lactic acid. In terms of energy yield, the conversion of pyruvic acid to either ethanol or lactic acid is very inefficient – only 2 moles of ATP are produced from every mole of sugar metabolised (compared with the potential 38 ATP from aerobic respiration). Fungi therefore need an abundant supply of sugars for growth in anaerobic conditions. We noted in Chapter 6 that fungi also need to be supplied with a wide range of other nutrients in anaerobic conditions, because the TCA cycle and several other reactions do not operate to provide the precursors for biosynthesis.
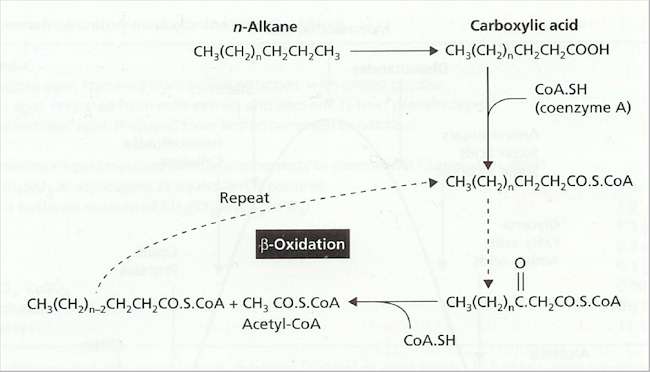
Fig. 7.4. Outline of the reactions in ß-oxidation – a process that occurs in the mitochondria of fungi. Long-chain fatty acids are activated by combining with coenzyme A and then enter a repeating cycle in which a molecule of acetyl-coenzyme A is removed in each turn of the cycle. Since most fatty acids have an even number of carbon atoms, this results in the complete conversion of fatty acids to acetyl-CoA. Long-chain hydrocarbons (n-alkanes; top left) such as those in aviation kerosene (Chapter 6) can also be processed through this pathway, but first they need to be oxidised by oxygenase enzymes, which catalyse the direct incorporation of molecular oxygen into the molecule.
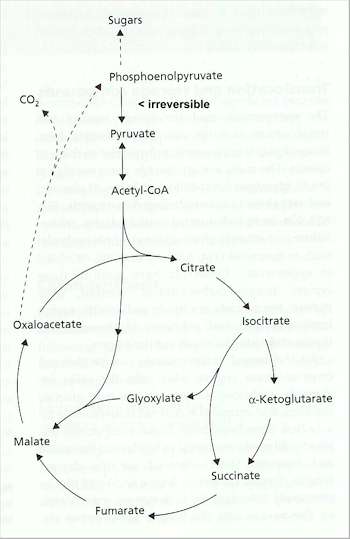
Fig. 7.5. Role of the glyoxylate cycle in generating sugars for biosynthesis when fungi are grown on non-sugar substrates such as acetate or organic acids.
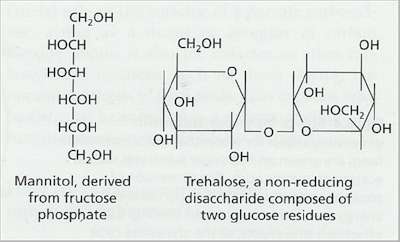
Fig. 7.6. The structure of mannitol and trehalose, two characteristic ‘fungal sugars’
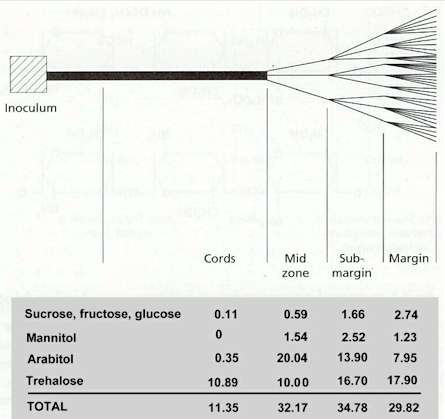
Fig. 7.7. Diagrammatic representation of the soluble carbohydrate contents in different regions of a colony of Serpula lacrymans growing across a sheet of Perspex from a precolonised wood block. Data are per cent of dry weight of the sugars in different zones. Based on data in C.Brownlee & D.H.Jennings (1981) Transactions of the British Mycological Society 77, 615-619.
Fig. 7.8. Part of a wooden door post heavily rotted by Serpula lacrymans. The surface of the timber is covered with dense yellow/white mycelial fronds of the fungus. The wood shows the characteristic block-like cracking (arrowheads) typical of dry rot. [© Jim Deacon]
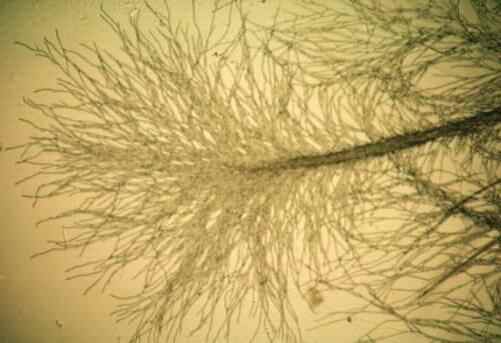
Fig. 7.9. Mycelial cords and fans of hyphae produced by a mycorrhizal fungus, Lactarius pubescens on an agar plate. [© Jim Deacon]
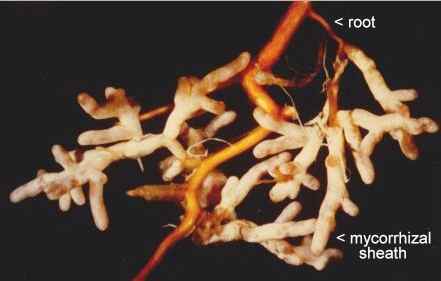
Fig. 7.10. Mycorrhizas of Amanita muscaria (the Fly Agaric) on a tree root system. The narrow roots are enclosed in a dense fungal sheath. Courtesy of Dr L.V.Fleming.
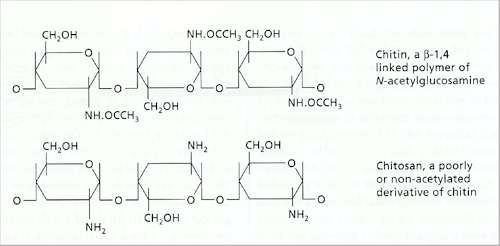
Fig. 7.11. Structure of chitin and its deacetylated derivative, chitosan |
|||||||||||