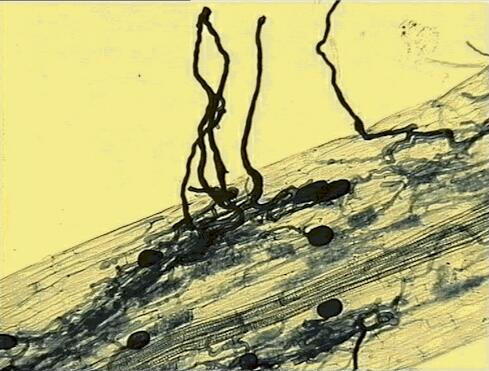
Environmental & Community Biology 1h MYCORRHIZAS: STUDY NOTES [To support Jim Deacon's lectures] What do all these things have in common? A mycorrhiza can be defined as a mutualistic association between a fungus and a root or other underground organ of a plant. Typically, the fungus penetrates the root tissues and obtains at least part of its carbon requirements from the plant, while the plant benefits by enhanced uptake of mineral nutrients via the fungal hyphae that grow through the soil. But there are exceptions, explained later. Mycorrhizal associations are found in more than 70% of vascular plants (angiosperms and gymnosperms) as well as many pteridophytes (ferns and their allies) and some bryophytes (especially liverworts). Indeed, structures resembling some of today's mycorrhizas have been found in plant fossils from the Devonian period. So, mycorrhizal associations developed early in the history of land plants, and have been maintained to this day. The relatively few non-mycorrhizal plants tend to grow in open habitats, where competition for soil nutrients may not be a major problem, but mycorrhizas are the norm in closed plant communities. Types of mycorrhiza There are several types of mycorrhiza, involving different types of plant and different types of fungus. The most widespread are the arbuscular mycorrhizas (AM), sometimes called vesicular-arbuscular mycorrhizas. These are found world-wide on many crop plants, wild herbaceous plants and trees, as well as on pteridophytes and some bryophytes. The fungi that form these mycorrhizas are members of the zygomycota, such as Glomus and Acaulospora spp. None of these can be grown in pure culture, away from their host plants. Roots appear quite normal when infected by these fungi, but staining of the root tissues reveals a substantial amount of internal colonisation (Figure 1).
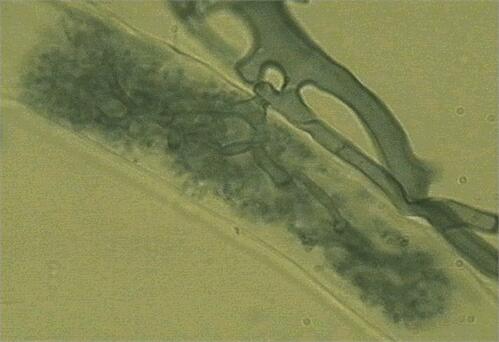
These fungi infect roots from spores or hyphal networks in soil, producing a characteristic appressorium-like structure on the root surface. From this, the hyphae grow between the root cortical cells and penetrate individual root cells to form arbuscules (finely branched tree-like structures) (Figure 2) and large vesicles within or between the root cells. The arbuscules are thought to be the sites of nutrient exchange, whereas the vesicles are probably storage bodies.
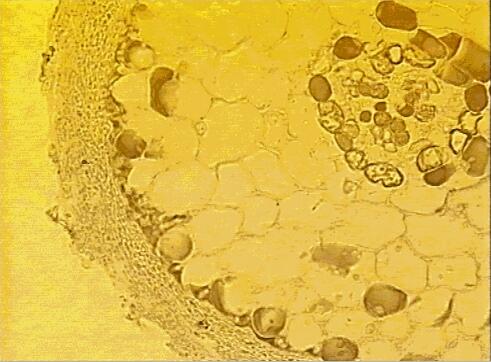
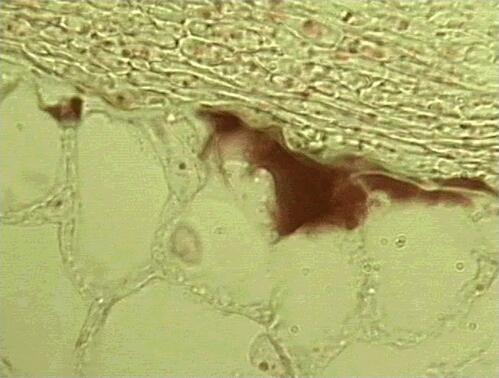
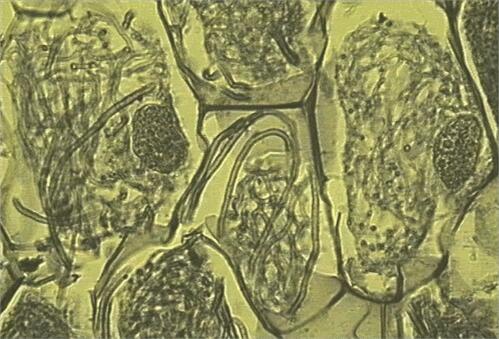
Many broad-leaved and coniferous trees of the temperate and boreal zones (e.g. pine, spruce, fir, oak, beech, birch, eucalypts) have ectotrophic mycorrhizas, or ectomycorrhizas, formed by fungi in the ascomycota and basidiomycota. The fruitbodies of these fungi include some of the common toadstools of the forest floor (e.g. Amanita, Boletus, Cortinarius, Hebeloma, Lactarius spp.) and also the truffles. The characteristic feature of ectomycorrhizas is that the fungal hyphae form a tissue-like sheath (mantle) on the surface of the nutrient-absorbing rootlets (Figs. 3, 4), and the rootlets themselves often are short and stumpy, lacking root hairs. From the sheath the hyphae extend into the soil, and also grow between the root cortical cells to form a 'Hartig net' (Fig. 5). The fungus is in close contact with the root cells in this region, but there is no penetration of the host cells, hence the term ectotrophic (outside-feeding) mycorrhiza.
Heathland plants in the family Ericaceae (Erica, Calluna, Vaccinium, etc.) have ericoid mycorrhizas, formed by fungi in the ascomycota and deuteromycota; for example Hymenoscyphus ericae. The hyphae of the fungus form coils within the root cells, and nutrient exchange is thought to occur primarily through these coils. Orchid mycorrhizas (Fig. 6) are different from any of those above, because the plant is actually parasitic on the fungus. Orchid seeds are extremely small, with minimal nutrient reserves, and they are triggered to germinate when they are infected by an appropriate fungus - typically a species of Rhizoctonia (basidiomycota). These fungi degrade cellulose and other components of soil organic matter. They infect the orchid cells to produce hyphal coils, but the coils are continuously digested and by this or other means the fungus provides the plant with a carbon source. Most orchids depend on this form of nutrition for their early growth because they may not produce chlorophyll for several months or years. A functionally similar type of mycorrhiza is found in the non-chlorophyllous Monotropa spp., but the fungi in these cases are either ectomycorrhizal species that are supplied with nutrients from a tree host (e.g. Boletus spp.) or they are tree pathogens such as Armillaria spp.
Why do plants have mycorrhizas? The orchid and monotropoid mycorrhizas are exceptional, but in all other cases the fungus obtains at least part of its carbon nutrition from the plant, while the extensive network of fungal hyphae in the soil provides an efficient means of absorbing mineral nutrients, some of which are then mobilised into the root cells.
Pea plants have been chemically mutated to be non-mycorrhizal - the mutants respond to attempted infection by a hypersensitive response similar to that against plant pathogens. This indicates that plants have had plenty of opportunity to become naturally immune to mycorrhizal infection, so mycorrhizas must confer an ecological advantage that offsets the cost. In the case of arbuscular mycorrhizas the major benefit is probably an enhanced uptake of phosphorus from soil. Even when soluble forms of phosphate fertiliser are added to cropping soils the phosphate ions rapidly form insoluble complexes with soil calcium or organic matter, so that the soluble pool of phosphate ion is quite low. In these circumstances, a non-mycorrhizal plant would need to have a continuously expanding root system, and continuous production of root hairs to tap new zones of soil. Fungal hyphae can replace this need, because they are thinner, extend further and have a much higher surface to volume ratio for mineral nutrient absorption. Ectomycorrhizas and ericoid mycorrhizas also are thought to facilitate nutrient uptake, but in these cases the uptake of nitrogen may be more important than of phosphorus. The reason is that the rates of mineralisation by breakdown of soil organic matter are quite low in cool, acidic soil conditions (the pH of many peatland sites is below 4.0) so that the release of nitrogen from organic matter can be a critical limiting factor for plant growth. Some of the fungi that form ericoid and ectomycorrhizas have been shown to degrade proteins in soil organic matter, obtaining a source of nitrogen that would otherwise be unavailable to plants. Mycorrhizas of many types have been shown to have other roles, such as protection of plants from attack by soil-borne pathogens, protection against toxic levels of trace elements, and enhancement of drought-tolerance because the fungal network can tap sources of water beyond the zone reached by the plant root system. Mycelial connections Most mycorrhizal fungi do not show a high degree of host-specificity, at least in artificial laboratory conditions. For example, a strain of arbuscular mycorrhizal fungus that infects tomato plants will also infect cotton or maple roots, and among the ectomycorrhizal fungi we also find 'generalists' with relatively wide host ranges on tree seedlings. This can mean that a hyphal network in soil might link several plants, even of different species (e.g. ectomycorrhizas formed by Laccaria spp., Hebeloma spp., Thelephora terrestris, Paxillus involutus). These mycelial links could have major significance in terms of competition or cooperation between plants in natural communities. We do not know the extent of this at present, but it has been shown experimentally that tree seedlings can be linked to a 'mother' tree or 'nurse' tree by a common , mycorrhizal network, such that 14CO2 fed to the leaves of a larger tree can be found as label in the roots and shoot tissues of nearby seedlings. Perhaps more important is the potential of the mycorrhizal network to retain and conserve mineral elements. An estimated 70-90% of ectomycorrhizal rootlets die and are replaced annually. If these rootlets were not interconnected by mycelia then they would be decomposed by the soil microbial community and at least some of the nutrients could be leached from the soil by rainwater. However, the mycorrhizal mycelial connections can help to retain mineral nutrients within the system, withdrawing them from the degenerating mycorrhizas to others that are still functioning. If this happens to any significant degree in mycorrhizal systems as a whole, then plants that are not tapped into the network would be at a significant disadvantage. The only available option would be to colonise disturbed habitats, and this is precisely where plants of the non-mycorrhizal families are commonly found. Most recently, another fascinating aspect of mycorrhizas has come to light (see the overview by David Read in Nature (November 1998) 396, 22-23; and the full paper on pp. 69-72 in the same issue). This work shows that the below-ground diversity of arbuscular mycorrhizal fungi is a major factor contributing to the above-ground plant biodiversity in ecosystems. In other words, if we wish to protect the above-ground diversity then we had better conserve the mycorrhizal fungal diversity in the soil! You can learn more about mycorrhizas by going to The Microbial World |