..
PROFILES OF FUNGI : RUST FUNGI (Basidiomycota) There are more than 4,000 species of rust fungi, found on a wide range of wild and cultivated plants in many parts of the world. All are members of the Basidiomycota (Order: Uredinales) but they are microscopic species, in contrast to the basidiomycota that produce the larger mushrooms and toadstools. Given such enormous diversity, it is difficult to do justice to this group, but we can begin by listing the major features of the "rusts" and then look more closely at some representative examples.
Fig 1. Part of a wheat leaf infected by Puccinia striiformis (yellow rust), showing numerous pustules of uredospores erupting through the leaf epidermis. [© Jim Deacon]
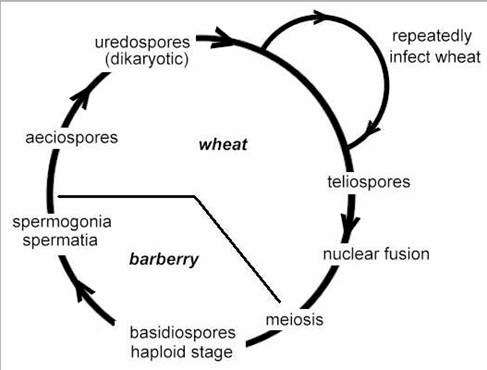
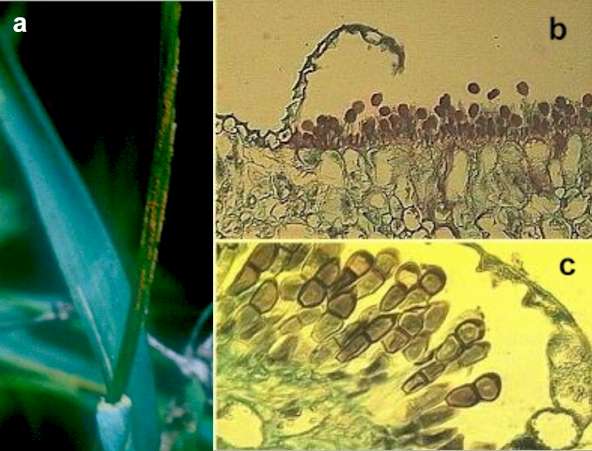
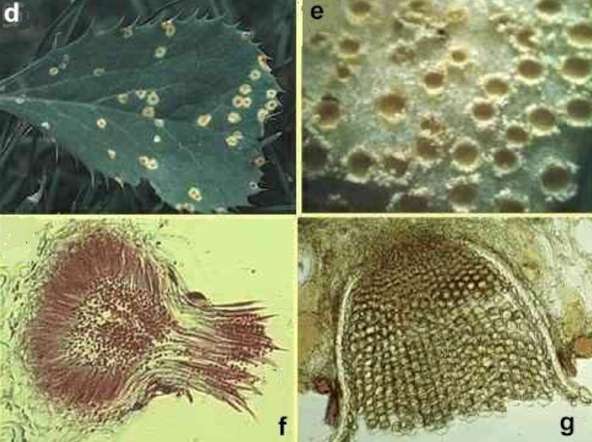
Black stem rust of wheat (Puccinia graminis) The disease cycle of black stem rust is shown in Fig. 3. We will begin at the dispersal phase, when rust pustules burst through the wheat stem in early spring and release large numbers of the characteristic rust-coloured uredospores, from structures called uredinia (Figs 3a and 3b). Each uredospore is single-celled but it contains two nuclei, one of each mating type, so the spores are dikaryotic as explained in Chapter 1. The uredospores are dispersed by wind and can undergo several cycles of infection in the course of a season, resulting in major epidemics. Towards the end of the season the production of uredospores ceases, and instead the pustules (now termed telia) produce 2-celled teliospores (Fig. 3c). These also are dikaryotic, with two nuclei in each cell, but the nuclei fuse to form diploid cells, and the fungus overwinters in this form. Early in the spring each cell of the teliospore germinates to a produce short mycelium. The diploid nucleus migrates into this and undergoes meiosis, leading to the production of 4 monokaryotic basidiospores (spores with only one haploid nucleus). These spores will only develop further if they land on a barberry leaf. Then they penetrate the leaf and, a few days later, produce flask-shaped spermogonia. These have receptive hyphae and also release many small spermatia (Figs 3d, 3f). Each spermogonium produces spermatia of a single mating type and they are exuded from the neck of the spermogonium in a sugary fluid. Flies and other insects are attracted to this, and when they visit a spermogonium of different mating type they transfer the spermatia. The spermatia of one mating type then fuse with receptive hyphae of the opposite mating type and transfer the nucleus through a pore in the wall. This nucleus then divides and migrates through the monokaryotic hyphae that grew from the basidiospores, producing a dikaryotic mycelium. By this time, the mycelium has grown through the thickness of the leaf and produces a further sporing structure that erupts through the lower epidermis – an aecium containing aeciospores (Figs 3e, 3g). The dikaryotic aeciospores can only infect wheat, and they give rise to infections on which uredospores will develop, thereby completing the cycle.
Fig 3 (above and below).. Stages in the life cycle of black stem rust of wheat caused by Puccinia graminis. Above: (a) Elongated stem lesions bearing uredospores. (b) Stained section of a wheat stem with a pustule of uredospores breaking though the plant epidermis. (c) Section of a pustule with teliospores. Below: d) Lesions containing spermogonia on the upper surface of a barberry leaf. (e) Aecia erupting through the lower epidermis of a barberry leaf. (f) A spermogonium, showing the tiny spermatia and receptive hyphae. (g) Cross-section of an aecium. [© Jim Deacon]
Other examples of rust fungi From the comments above it should be clear that rust fungi are extremely common, even on wild plants, and they often live in balance with their hosts. But this balance is upset when susceptible host plants are grown in monoculture over large acreages, using single "pure-bred" cultivars that are genetically uniform. These are the conditions that create disease epidemics. To end this account, the following images illustrate some of the diversity of the common rust fungi.
Fig 4. Left: Rust pustules on the stem of a common weed, groundsel (Senecio vulgaris). Right: Cup-shaped aecia on a Senecio leaf. [© Jim Deacon]
Fig 5. Rust of stinging nettle (Urtica dioica) caused by Puccinia caricina. The infection resulted in a 3 cm zone of deformed growth on the stem, but with no other significant effect. The aecia are clearly seen. The primary host of this rust is Carex spp. (sedges) on which the uredospores and and pycnia are produced. [© Jim Deacon]
Fig 6. A rust canker on the stem of a rose bush. Note the large mass of uredospores on the leaves and young shoots, which soon wither and die. The rest of the canes showed no ill effect. [© Jim Deacon]
Fig 7. Left: Black pustules on the underside of a a blackberry leaf (Rubus fruticosus), caused by the rust fungus Phragmidium violaceum. Right: Teliospores of P. violaceum scraped from one of the black pustules. Each spore consists of a stalk and a short chain of usually 4-5 darkly pigmented cells. [© Jim Deacon] |